

Struktura uracylu, funkcje, właściwości, synteza
- 3125
- 309
- Filip Augustyn
On Uracyl Jest to podstawa azotu pirymidyny, która występuje w kwasie rybonukleinowym (RNA). Jest to jedna z cech, które odróżnia RNA od kwasu deoksyrybonukleinowego (DNA), ponieważ ta ostatnia ma Timinę zamiast uracylu. Obie substancje, uracyl i Timina, różnią się tylko, że druga ma grupę metylową.
Z ewolucyjnego punktu widzenia zaproponowano, że RNA był pierwszą cząsteczką, która przechowywała informacje genetyczne i pracowała jako katalizator w komórkach, przed DNA i enzymami. Z tego powodu uważa się, że Uracil odgrywał kluczową rolę w ewolucji życia.
Źródło: Kemikungen [domena publiczna] W żywych istotach uracyl nie występuje swobodnie, ale zwykle nukleotydy monofosforanowe (UMP), difosforan (UDP) i trfosforan (UTP). Te nukleotydy uracylu mają różne funkcje, takie jak biosynteza RNA i glikogen, izomeryczna interkonwersja cukrów i regulacja syntazy syntaminy.
[TOC]
Struktura i właściwości
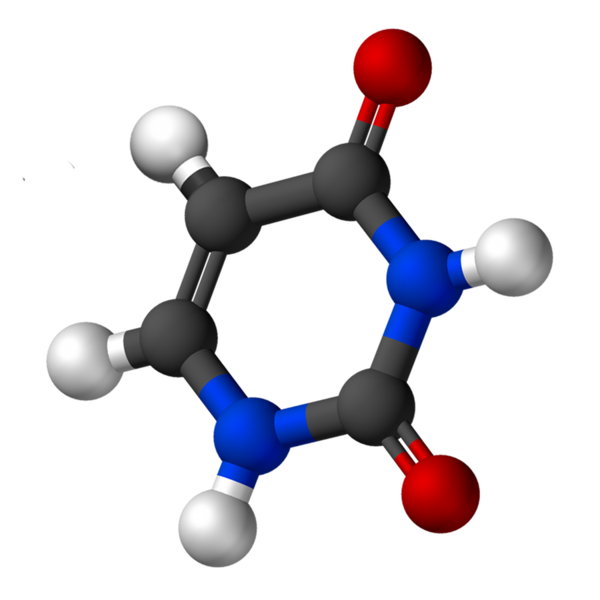
Uracyl, zwany 2.4-dioksypirydyny, ma wzór empiryczny C4H4N2ALBO2, którego masa cząsteczkowa wynosi 112,09 g/mol i jest oczyszczona jako biały proszek.
Struktura urydyny jest pierścieniem heterocyklicznym z czterema atomami węgla i dwoma azotem, z podwójnymi alternatywnymi wiązaniami. To jest płaskie.
Ma rozpuszczalność 50 mg/ml, 25 ° C, w 1M wodorotlenku sodu i PKA między 7.9 i 8.2. Długość fali, w której występuje maksymalna absorbancja (ʎMax) Jest to od 258 do 260 nm.
Biosynteza
Istnieje wspólna ścieżka biosyntezy nukleotydów pirymidynowych (uracyl i cytokina). Pierwszym krokiem jest biosynteza fosforanu karbamowskiego z CO2 i NH4+, który jest katalizowany przez fosforan syntazy karbamowskiej.
Pirymidyna jest zbudowana z fosforanu węglowodanów i asparaginianu. Obie substancje reagują i tworzą N-karbamoilaspartato, reakcja katalizowana przez asparagan Transcabamoilasa (ATCASA). Zamknięcie pierścienia pirymidynowego można odwodnić przez dihydrootazę i wytwarza di-dihydrotatatatoan.
L-dihydrotoate jest utleniany i przekształcany w Orotan; Akceptor elektronów to NAD+. Jest to reakcja katalizowana przez dehydrogenazę dihydroorotatanową. Kolejny krok polega na przeniesieniu grupy fosforebozylowej, od pirofosforanu fosforytylowego (PRPP), do orotatu. Postać gradidilato (WPO) i piroffosfat nieorganiczny (PPI), katalizowany przez transforyzę fosforabosyl.
Ostatni krok obejmuje dekarboksylację pierścienia pirymidynowego grupy (WPO). Postać Uridilaato (monofosforan urydyny-5', UMP), który jest katalizowany przez dekarboksylazę.
Następnie, poprzez udział kinazy, grupa fosforanowa jest przenoszona z ATP do UMP, tworząc UDP (diphosforan urydyny-5'). Ten ostatni jest powtarzany, tworząc UTP (Uridin-5'-triffosforan).
Może ci służyć: gałęzie biologii i czego się ucząRegulacja biosyntezy
W bakteriach regulacja biosyntezy pirymidyny zachodzi poprzez ujemne sprzężenie zwrotne, na poziomie asparaganu Transcabamoilasa (ATCASA).
Ten enzym jest hamowany przez CTP (cytidin-5'-trofosforan), który jest końcowym produktem szlaku biosyntezy pirymidynów. Atcas.
U zwierząt regulacja biosyntezy pirymidyny zachodzi poprzez ujemne sprzężenie zwrotne, na poziomie dwóch enzymów: 1) fosforanu syntazy karbamowskiej II, który jest hamowany przez UTP i aktywowany przez ATP i PRPP; oraz 2) OMP Discarbokslaza, która jest hamowana przez iloczyn reakcji, którą sama katalizuje, UMP. Prędkość biosynteza OMP zmienia się w zależności od dostępności PRPP.
Funkcja w biosyntezy RNA
Uracyl jest obecny we wszystkich typach RNA, takich jak Messenger RNA (MNA), RNA transferowy (ARNT) i rybosomalny RNA (RNA). Biosynteza tych cząsteczek odbywa się w procesie zwanym transkrypcją.
Podczas transkrypcji informacje zawarte w DNA są kopiowane w RNA przez RNA polimerazy Pon. Proces odwrotny, w którym informacje zawarte w RNA są kopiowane w DNA, odbywa się w niektórych wirusach i roślinach przez odwrotną transkryptazę.
Biosynteza RNA wymaga nukleozydów tryposforanowych (NTP), a mianowicie: tryfosforan urydyna (UTP), cytydyny tryposforanu (CTP), adenine tryposforan adenina (ATP) i tryfosforan guanine (GTP). Reakcja to:
(RNA)n marnotrawstwo + NTP -> (RNA)N+1 Waste + PPI
Hydroliza pirofosforanu nieorganicznego (PPI) zapewnia energię do biosyntezy RNA.
Funkcja w biosyntezy cukru
Estry cukrowe są bardzo powszechne w żywych organizmach. Niektóre z tych estrów są dyfosfami estrów nukleozydów, takie jak UDP-SUGAR, które są bardzo obfite w komórkach. UDP-SUGAR obejmuje biosyntezę disacharydów, oligosacharydów i polisacharydów.
W roślinach biosynteza sacharozy odbywa się na dwa sposoby: główny i wtórny.
Główna trasa polega na przeniesieniu d-glukozy z UDP-D-glukozy do fruceazy D w celu utworzenia sacharozy i UDP. Druga droga zawiera dwa etapy: zaczyna się od UDP-D-glukozy i fruktozy-6-fosforanu i kulminacją z tworzeniem się sacharozy i fosforanu.
W gruczołach sutkowych biosynteza laktozy odbywa się z UDP-D-galaktozy i glukozy.
Może ci służyć: oddziały biochemiiW roślinach biosynteza celulozy przeprowadza się przez ciągłą kondensa. Podobnie biosynteza amylozy i amylopektyny wymaga UDP-glukozy jako podłoża dawcy glukozy do rosnącego łańcucha.
U zwierząt zarówno UDP-glukoza, jak i ADP-glukoza są stosowane do biosyntezy glikogenu. Podobnie biosynteza siarczanu chondroitiny wymaga UDP-ksylozy, UDP-galaktozy i UDP-glukuronato.
Funkcja w izomerycznej interkonwersji cukrów
Konwersja galaktozy do pośrednika glikolizy odbywa się przez Leloir Road. Jeden z etapów tej trasy jest katalizowany przez enzym UDP-galaktozę-4-epimerazę, który ułatwia interpretację UDP-galaktozy do UDP-glukozy.
Funkcja w biosyntezy glikoprotein
Podczas biosyntezy glikoprotein białka przechodzą przez worki CIS, pożywki i trans aparatu Golgiego.
Każda z tych toreb ma zestaw enzymów, które przetwarzają glikoproteiny. Monomery cukrowe, takie jak glukoza i galaktoza.
Nukleotydy-heksosę są transportowane do zbiorników Golgiego przez antyportę. UDP-galaktoza (UDP-GAL) i UDP-N-acetylgalaktosamina (UDP-galnac) wchodzą do zbiorników z cytosolu za pomocą wymiany przez UMP.
W zbiorniku Golgiego fosfataza hydrolizuje grupę fosforanową postać UDP i UMP i PI. UDP pochodzi z reakcji katalizowanych przez galaktozylotransferazę i N-acetylgalaktosamiltransferazę. UMP utworzone przez fosfatazę służy do wymiany nukleotydów-heksozy.
Funkcja w regulacji syntazy
Mechanizm regulacji syntazy glutaminy jest kowalencyjna modyfikacja, która składa się z adenilacji, która nieaktywna i flanka, która ją aktywuje. Ta kowalencyjna modyfikacja jest odwracalna i katalizowana przez adenylotransferazę.
Aktywność adenylotransferazy jest modulowana przez połączenie białka PII, które jest regulowane przez kowalencyjną modyfikację, urydynację.
Zarówno Uridililacja, jak i przesunięcie są przeprowadzane przez Uridiltransferazę. W tym enzymie aktywność urydylacji jest spowodowana glutaminą i fosforanem i jest aktywowana przez związek alfa-zotoglutaranu i ATP do PII.
Funkcja w wydaniu RNA
Niektóre RNM są edytowane przed tłumaczeniem. W niektórych organizmach eukariotycznych, takich jak Trypanosoma Brucei, Istnieje wydanie RNA transkrypcji genu podjednostki II oksydazy cytochromu. Dzieje się tak poprzez wstawienie odpadów uracylowych, reakcji katalizowanej przez końcową uridiltransferazę.
Może ci służyć: curl: kompozycja, części, znaczeniePrzewodnik RNA, uzupełniający edytowany produkt, działa równie umiarkowane w procesie edycji. Pary zasad utworzone między początkowym transkryptem a przewodnikiem RNA implikuje pary zasad g = u, które nie są rekordowe i są powszechne w RNA.
Biosynteza UDP-glukozy
W warunkach fizjologicznych biosynteza glikogenu z fosforanu glukozy-1 jest termodynamicznie niemożliwa (dodatnie ΔG). Z tego powodu przed biosyntezą następuje aktywacja glukozy-1-fosforanu (G1P). Ten G1P i UTP łączą reakcję z utworzeniem glukozy difosforanu urydyny (UDP-glukoza lub UDPG).
Reakcja jest katalizowana przez pirofosforylazę UDP-glukozę i jest następująca:
G1P + UTP -> UDP -Glucosa + 2pi.
Gibbs Wariacja energii swobodnej w tym etapie jest duża i ujemna (-33,5 kJ/mol). Podczas reakcji tlenu G1P atakuje atom Alforo Alpha Phosforus w UDP-glukozy i pirofosforanu nieorganicznego (PPI). Następnie PPI jest hydrolizowany przez nieorganiczne pirofosforany, których energia hydrolizy jest tym, która napędza ogólną reakcję.
UDP-glukoza jest substancją „wysokiej energii”. Pozwala tworzyć wiązania glikozydowe między pozostałością glukozy a rosnącym łańcuchem polisacharydowym. Ta sama zasada energii ma zastosowanie do reakcji, w których uczestniczy UDP-SUGAR, takie jak biosynteza disacharydów, oligosacharydów i glikoprotein.
Uracyl DNA glikosilasa
Istnieją zmiany DNA, które występują spontanicznie. Jedną z tych zmian jest sprontane. W takim przypadku naprawa odbywa się z powodu zmodyfikowanej podstawy DNA za pomocą enzymu zwanego uracyl DNA glikosilasa.
Glikosilasa DNA enzymu glikosilasa eliminuje uszkodzoną cytokinę (uracyl), wytwarzając resztę dezoksyrybozy, której brakuje podstawy azotu, zwana miejscem AP (miejsce apurynowe-apirymidyniczne).
Następnie endonukleaza enzym AP wykonuje cięcie w szkielecie fosfodiestrowym w miejscu AP, eliminując pozostałości fosforanowe cukru. Polimeraza DNA I przywraca uszkodzoną pasmę.
Bibliografia
- Bohinski, r. 1991. Biochemia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biochemia. Redakcja Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, s. 1., Baltimore, zm., Darnell, J. 2003. Biologia komórkowa i molekularna. Redakcja Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madryt, Meksyk, Sāo Paulo.
- Nelson, zm. L., Cox, m. M. 2008. Lehninger-Principles of Biochemistry. W.H. Freeman, Nowy Jork.
- Voet, d. i Voet, J. 2004. Biochemia. John Wiley and Sons, USA.