

Sfingolipidy, czym są, cechy, funkcje, synteza
- 1832
- 291
- Herbert Wróblewski
Co to są phylingolipids?
Sphingolipids Reprezentują jedną z trzech wielkich rodzin lipidów obecnych w błonach biologicznych. Podobnie jak glyceofosfolipidy i sterole, są to cząsteczki amfipatyczne o hydrofilowym regionie polarnym i hydrofobowym obszarze apolowym.
Zostały po raz pierwszy opisane w 1884 roku przez Johanna L. W. Thudichum, który opisał trzy sfingolipidy (sfhingomyeline, mózg i mózg -mózg), które należą do trzech znanych klas: fosfosfosfingolipids, glikosfingolipidy neutralne i kwaśne.
W przeciwieństwie do gliceofosfolipidów, sphyingolipidy nie są zbudowane na 3-fosforanowej cząsteczce glicerolu jako główny szkielet, ale są związkami pochodzącymi z Sfiny, aminoalkoholu z długim łańcuchem węglowodorowym przyłączonym przez wiązanie amida.
IStruktura sfingolipidów
Wszystkie sfingolipidy pochodzą z serii L, która jest skondensowana długim łańcuchem kwasu tłuszczowego z tworzeniem zasady sfingoidalnej, znanej również jako podstawa długiego łańcucha (LCB).
Najczęstszymi zasadami są sfinganina i sfinksyna, które różnią się od siebie w obecności podwójnego wiązania trans Między węgle 4 i 5 kwasu tłuszczowego sfinksyny.
Węglowodory 1, 2 i 3 sferksyny są strukturalnie analogiczne do glicerolu glicerofosfolipidów glicerol. Kiedy wiązania amida wiąże kwas tłuszczowy z węglem 2 sphin.
Kwasy tłuszczowe długiego łańcucha, które stanowią hydrofobowe regiony tych lipidów, mogą być bardzo zróżnicowane. Długości wahają się od 14 do 22 atomów węgla, które mogą mieć różne stopnie nasycenia, zwykle między węgliami 4 i 5.
W pozycjach 4 lub 6 mogą mieć grupy wodorotlenowe i podwójne wiązania w innych pozycjach, a nawet konsekwencje, takie jak grupy metylowe.
Charakterystyka
Łańcuchy kwasów tłuszczowych połączone wiązaniami amidy z ceramidami są powszechnie nasycone i mają tendencję do większej długości niż te stwierdzone w glicerofosfolipidach, co wydaje się być kluczowe dla ich aktywności biologicznej ich.
Charakterystyczną cechą szkieletu sfingolipidowego jest to, że mogą mieć dodatnie obciążenie netto do neutralnego pH, dziwne wśród cząsteczek lipidowych.
Jednakże PkA grupy aminowej jest niski w odniesieniu do prostej aminy, między 7 a 8, więc część cząsteczki nie jest ładowana do fizjologicznego pH, co może wyjaśnić ich „swobodny” ruch wśród Bilapas.
Tradycyjna klasyfikacja sfingolipidów wynika z wielokrotnych modyfikacji, które może ponieść cząsteczka ceramidowa, szczególnie pod względem podstawień grup głównych polarnych.
Może ci służyć: Hepadnavirus: Charakterystyka, morfologia, leczenieFunkcje
Sfingolipidy są niezbędne u zwierząt, roślin i grzybów, a także w niektórych organizmach i wirusach prokariotycznych.
-Funkcje strukturalne
Sfingolipidy modulują właściwości fizyczne membran, w tym ich płynność, grubość i krzywiznę. Modułowe właściwości te dają im również bezpośredni wpływ na przestrzenną organizację białek błonowych.
W lipidowych „tratwach”
W błonach biologicznych dynamiczne mikro domeny można wykryć z niższą płynnością utworzoną przez cząsteczki cholesterolu i sfingolipidów zwane balsami lipidowymi.
Struktury te występują naturalnie i utrzymują ścisły związek z kompleksowymi białkami, receptorami powierzchniowymi komórkami i białkami sygnalizacyjnymi, transporterami i innymi białkami z kotwicami glikozylofosfatydyloinitolu (GPI) (GPI) (GPI).
-Funkcje sygnalizacyjne
Mają funkcje, takie jak cząsteczki sygnalizacyjne, które działają jako drugie posłańcy lub jako wydzielane ligandy dla receptorów na powierzchni komórki.
Jako wtórni posłańcy mogą uczestniczyć w regulacji homeostazy wapnia, wzrostu komórek, nowotworu i supresji apoptozy. Ponadto aktywność wielu integralnych i obwodowych białek błonowych zależy od jego związku z sfingolipidami.
Wiele interakcji międzykomórkowych i komórkowych z otoczeniem zależy od ekspozycji różnych grup polarnych sfingolipidów w kierunku zewnętrznej powierzchni błony plazmatycznej.
Związek glikosfingolipidów i lektyn ma kluczowe znaczenie dla powiązania mieliny z aksonami, przyczepności neutrofili do śródbłonka itp.
Przez -produkty jego metabolizmu
Najważniejszymi sfingolipidami sygnalizacyjnymi są zasady długiego łańcucha lub sfinkses i ceramidy, a także ich fosforylowane pochodne, takie jak 1-fosforanowa sPhygozyna.
Produkty metabolizmu wielu sfingolipidów aktywują lub hamują wiele białych dół (białka kinazy, fosfatoza i inne), które kontrolują zachowania komórkowe tak złożone jak wzrost, różnicowanie i apoptoza.
-Jako receptory błony
Niektóre patogeny używają GLoysphingolipidów jako receptorów w celu pośredniczenia w ich wejściu do komórek gospodarza lub dostarczania im czynników wirulencji.
Wykazano, że sphyglyipidy uczestniczą w wielu zdarzeniach komórkowych, takich jak wydzielanie, endocytoza, chemiotaksja, neurotransmisja, angiogeneza i zapalenie.
Są również zaangażowane w ruch błony, więc wpływają na internalizację receptorów, porządku, ruchu i fuzji pęcherzyków wydzielniczych w odpowiedzi na różne bodźce.
Grupy sfingolipidów
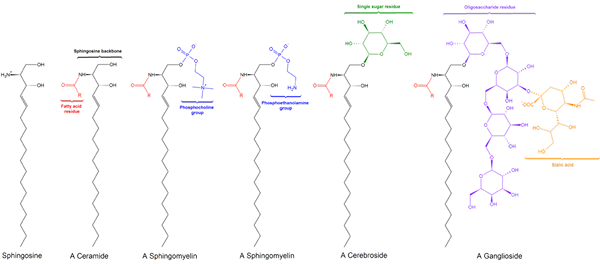
Istnieją trzy podklasy sfingolipidów, wszystkie pochodzące z ceramidu i które różnią się od siebie grupami polarnymi, a mianowicie: phylingomyelins, glikolipidy i zwojów.
SphingomyieLinas
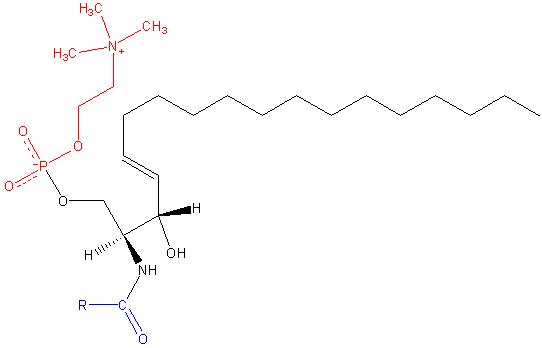 Sphingomilein. Czarny: sfinksyna. Czerwony: fosfokolina. Niebieski: kwas tłuszczowy.
Sphingomilein. Czarny: sfinksyna. Czerwony: fosfokolina. Niebieski: kwas tłuszczowy. Zawierają one fosfokolinę lub fosfoetanoloaminę jako grupę polarną, więc są klasyfikowane jako fosfolipidy wraz z glyceofosfolipidami. Wyglądają oczywiście, fosfatydylokolinom w trójwymiarowej strukturze i właściwościach ogólnych, ponieważ nie mają opłat w swoich głowach polarnych.
Może ci służyć: enolaza: struktura, mechanizm działania, funkcjeSą obecne w błonach plazmatycznych komórek zwierzęcy.
Glikolipidy lub neutralne glikosfingolipidy (bez obciążenia)
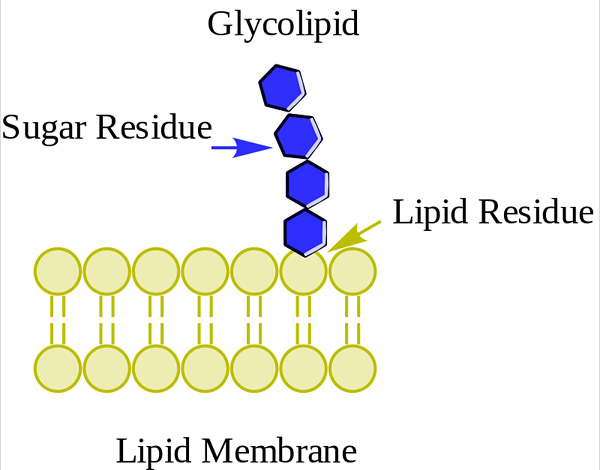 Glucoolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)], z Wikimedia Commons
Glucoolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)], z Wikimedia Commons Są przede wszystkim na zewnętrznej powierzchni błony plazmatycznej i mają jeden lub więcej cukrów jako grupa polarna bezpośrednio zjednoczona z hydroksylami węglową części ceramidowej. Nie mają grup fosforanowych. Ponieważ pH 7 nie ma obciążenia, nazywane są neutralnymi glikolipidami.
Cerebroozydy mają pojedynczą cząsteczkę cukru połączoną z ceramidą. Osoby zawierające galaktozę znajdują się w błonach plazmatycznych komórek tkankowych innych niż nerwy. Globozydy to GoustoEsPhingolipids z dwoma lub więcej cukrami, normalnie D-glikozą, D-galaktozy lub N-acetylo-D-galaktozaminy.
Gangliósydos lub glikosfingolipidy
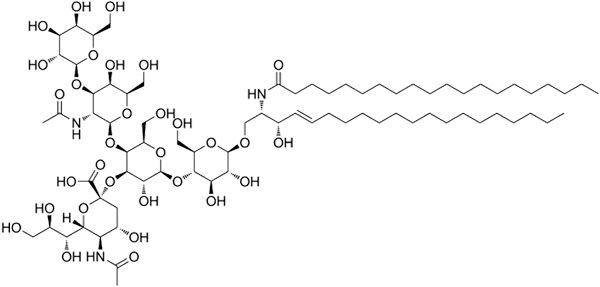 Struktura ganglozydowa GM1
Struktura ganglozydowa GM1 To są najbardziej złożone sfingolipidy. Mają oligosacharydy jako grupę polarną i jedną lub więcej końcowych reszt kwasu N-acetylamuraminowego, zwane także kwasem Siral. Kwas siaminowy wnosi do zwojów obciążenie ujemne do pH 7, co odróżnia ich od neutralnych glikosfingolipidów.
Nomenklatura tego rodzaju sfingolipidów zależy od ilości odpadów kwasu sinalicznego obecnego w części oligosacharydowej głowicy polarnej.
Synteza
Długa klimatyzacja lub sfinomowa cząsteczka zasady jest syntetyzowana w retikulum endoplazmatycznym (ER), a dodanie grupy polarnej na czele tych lipidów występuje później w kompleksie Golgiego. U ssaków pewna synteza sfingolipidów może również wystąpić w mitochondriach.
Po zakończeniu syntezy w kompleksie Golgiego, sfingolipidy są transportowane do innych przedziałów komórkowych za pomocą mechanizmów za pośrednictwem pęcherzyków.
Biosynteza sfingolipidów składa się z trzech fundamentalnych zdarzeń: syntezy zasad długiego łańcucha, biosyntezy ceramidów przez połączenie kwasu tłuszczowego poprzez wiązanie amidy, a na koniec tworzenie złożonych sfingolipidów za pomocą związku grup polarnych w węglku 1 podstawy sfingoide.
Oprócz syntezy Novo, Sfingolipidy mogą również tworzyć się przez wymianę lub recykling podstaw i ceramid długiego łańcucha, które mogą zasilać basen sfingolipidów.
Synteza szkieletu ceramidu
Biosynteza ceramidy, szkielet sfingolipidów, zaczyna się od dekarboksylicyjnej kondensacji cząsteczki palmitail-CoA. Reakcję jest katalizowana przez heterodimeryczną, zależną od fosforanu pirydoksalnego serynowego (SPT).
Może ci służyć: mikrobiota: skład, funkcje i gdzie to jestTen enzym jest hamowany przez β-faal-l-alaniny i L-ckloseryny. U drożdży jest kodowany przez dwa geny, podczas gdy u ssaków istnieją trzy geny dla tego enzymu. Aktywne miejsce znajduje się w cytoplazmatycznej powierzchni retikulum endoplazmatycznego.
Rola tego pierwszego enzymu jest zachowana we wszystkich badanych organizmach. Istnieją jednak pewne różnice między taksonami, które mają związek z podkomórkową lokalizacją enzymu: bakterie to cytoplazmatyczne, drożdże, rośliny i zwierzęta są w retikulum endoplazmatycznym.
3-Zoesfinganina jest następnie zmniejszona przez zależne od NADPH 3-ZO-KETOSFINGANINYNY. Syntaza dihydroceramidowa (transferaza n-acylowa sfinganina) acetyla, a następnie sfinanina do wytworzenia dihydroceramid. Ceramid jest następnie tworzony przez dihydroceramid deaturazę/reduktazę, która wstawia podwójne wiązanie trans W pozycji 4-5.
U ssaków istnieje wiele syntazów ceramidowych, z których każda łączy określony łańcuch kwasu tłuszczowego z zasadami długich łańcuchów. Dlatego ceramidy syntezy i inne enzymy, elongasas, stanowią główne źródło różnorodności kwasów tłuszczowych w sfingolipidach.
Metabolizm
Degradacja sfingolipidów odbywa się przez glikohydrole i enzymy sfingomyelinazy, które są odpowiedzialne za usunięcie modyfikacji grup polarnych. Z drugiej strony ceramidazy regenerują długie podstawy łańcucha z ceramid.
Zwoje są degradowane przez zestaw enzymów lizosomalnych, które katalizują krok po kroku jednostek cukru, wytwarzając ceramid w końcu.
Innym sposobem degradacji jest internaliza.
Nie wszystkie długie podstawy łańcucha są poddawane recyklingowi, endoplazmatyczne zachowują. Ten mechanizm degradacji obejmuje fosforylację zamiast acylacji LCB, powodując powstanie cząsteczek sygnalizacyjnych, które mogą być rozpuszczalnym substratami dla enzymu Liases.
Rozporządzenie
Metabolizm tych lipidów jest regulowany na kilku poziomach, jednym z nich jest metal enzymów odpowiedzialny za syntezę, ich modyfikacje po translacyjnych i te same mechanizmy tego samego.
Niektóre mechanizmy regulacji są specyfikacjami komórek, albo w celu kontrolowania czasu rozwoju komórek, w którym są one wytwarzane lub w odpowiedzi na określone sygnały.
Bibliografia
- Bartke, n., & Hannun i. (2009). Bioaktywne szarpnięcia: metabolizm i funkcja. Journal of Lipid Research, pięćdziesiąt, 19.
- Breslow, d. K. (2013). Homeostaza sfingolipidów w retikulum endoplazmatycznym i nie tylko. Perspektywy Cold Spring Harbor w biologii, 5(4), A013326.