

Struktura kwasów tłuszczowych, typy, funkcje, biosynteza
- 3603
- 442
- Marianna Czarnecki
Kwasy tłuszczowe Są to organiczne makrocząsteczki pochodzące z węglowodorów, które składają się z długich łańcuchów atomów węgla i wodoru, które są hydrofobowe (są rozpuszczalne w tłuszczu) i są podstawą strukturalną tłuszczów i lipidów.
Są to bardzo zróżnicowane cząsteczki, które odróżniają się od siebie przez długość łańcuchów węglowodorów i obecność, liczbę, pozycję i/lub konfigurację podwójnych wiązań.
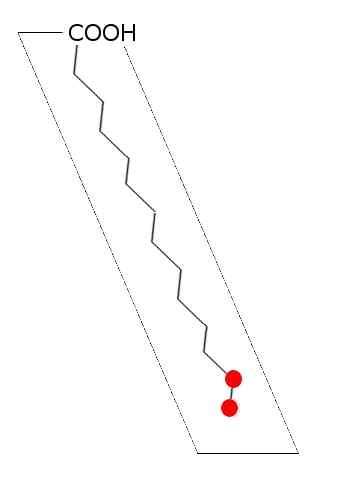
 Ogólny schemat nasyconego kwasu tłuszczowego (źródło: laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licencje/by-sa/3.0/)] przez Wikimedia Commons)
Ogólny schemat nasyconego kwasu tłuszczowego (źródło: laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licencje/by-sa/3.0/)] przez Wikimedia Commons) W lipidach zwierząt, roślin, grzybów i mikroorganizmów, takich jak bakterie i drożdże, opisano ponad 100 rodzajów różnych kwasów tłuszczowych i są one uważane za specyficzne gatunki i tkankę u większości żywych istot.
Oleje i tłuszcze, które człowiek spożywa codziennie, są pochodzenia zwierzęcego lub rośliny, składają się głównie z kwasów tłuszczowych.
[TOC]
Wstęp
 Masło między innymi składa się zasadniczo z kwasów tłuszczowych (źródło: Africa Studio, przez Pixabay.com)
Masło między innymi składa się zasadniczo z kwasów tłuszczowych (źródło: Africa Studio, przez Pixabay.com) Cząsteczki kwasu tłuszczowego pełnią ważne funkcje na poziomie komórkowym, co czyni je niezbędnymi składnikami, a ponieważ niektórych z nich nie mogą być syntetyzowane przez zwierzęta, muszą je uzyskać z diety.
Kwasy tłuszczowe są rzadkie jako wolne gatunki w cytosolu komórkowym, więc na ogół są częścią innych koniugatów molekularnych, takich jak:
- Lipidy w błonach biologicznych.
- Trójglicerydy lub estry kwasów tłuszczowych, które służą jako rezerwat u roślin i zwierząt.
- Woski, które są stałymi estrami o długim łańcuchu kwasów tłuszczowych i alkoholi.
- Inne podobne substancje.
U zwierząt kwasy tłuszczowe są przechowywane w cytoplazmie komórek jako małe krople tłuszczu złożone z kompleksu zwanego triacyloglicerolem, który jest niczym więcej niż cząsteczką glicerolu, do której połączyła się w każdym z jego atomów węgla, łańcucha tłuszczowego kwasu przez wiązania typu estra.
Podczas gdy bakterie mają krótkie i zwykle jednonienasycone kwasy tłuszczowe, w naturze jest to powszechne.
Struktura
Kwasy tłuszczowe to cząsteczki amfipatyczne, to znaczy mają dwa chemicznie zdefiniowane regiony: hydrofilowy obszar polarny i hydrofobowy obszar apolowy.
Region hydrofobowy składa się z długiego łańcucha węglowodorów, który w kategoriach chemicznych nie jest zbyt reaktywny. Z drugiej strony region hydrofilowy składa się z terminali grupy karboksylowej (-COH), która zachowuje się jak kwas.
Ta końcowa grupa karboksylowa lub kwas karboksylowy jest jonizowany w roztworze, jest wysoce reaktywna (chemicznie) i jest bardzo hydrofilowa, więc reprezentuje kowalencyjne miejsce połączenia między kwasem tłuszczowym a innymi cząsteczkami.
Długość łańcuchów węglowodorów kwasów tłuszczowych zwykle ma parzystą liczbę atomów węgla, co jest ściśle związane z procesem biosyntezy, według którego są one wytwarzane, ponieważ ich wzrost występuje z par węgla.
Najczęstsze kwasy tłuszczowe mają łańcuchy od 16 do 18 atomów węgla i zwierząt, łańcuchy te nie są rozgałęzione.
Klasyfikacja
Kwasy tłuszczowe są podzielone na dwie duże grupy zgodnie z naturą łączy, które je komponują, to znaczy, zgodnie z obecnością prostych wiązań lub podwójnych wiązań między atomami węgla ich łańcuchów węglowodorowych.
Zatem istnieją nasycone i nienasycone kwasy tłuszczowe.
- Kwasy tłuszczowe nasycone mają tylko proste wiązania węgla i węgla, a wszystkie ich atomy węgla są „nasycone” lub powiązane z cząsteczkami wodoru.
- Nienasycone kwasy tłuszczowe mają jeden lub więcej wiązań węgla z podwójnym węglem i nie wszystkie z nich są przyłączone do atomu wodoru.
Nienasycone kwasy tłuszczowe są również podzielone zgodnie z liczbą nienasycenia (podwójne wiązania) u jednoszsynowanych, z pojedynczym podwójnym wiązaniem i wielonienasyconym, które mają więcej niż jeden.
Nasycone kwasy tłuszczowe
Zwykle mają od 4 do 26 atomów węgla zjednoczonych przez proste linki. Jego temperatura topnienia jest wprost proporcjonalna do długości łańcucha, to znaczy do jego masy cząsteczkowej.
Kwasy tłuszczowe, które mają od 4 do 8 węgli, są ciekłe w temperaturze 25 ° C i są tymi, które tworzą oleje jadalne, podczas gdy te z ponad 10 atomami węgla są stałe.
Jednym z najczęstszych jest kwas laurowy, który jest obfity w nasionach palm i kokosu; Kwas palmitynowy, który występuje w palmie, kakao i masło świń, a kwas stearynowy, który znajduje się w kakao i olejach uwodornionych.
Są to kwasy tłuszczowe o znacznie większej stabilności niż nienasycone kwasy tłuszczowe, szczególnie w obliczu utleniania, przynajmniej w warunkach fizjologicznych.
Dzięki faktowi, że proste wiązania węgla - węglowe mogą obracać się za darmo.
Nienaturowane kwasy tłuszczowe
Te kwasy tłuszczowe są wysoce reaktywne i podatne na nasycenie i utlenianie. Są powszechne w roślinach i organizmach morskich. Ci, którzy mają pojedynczą podwójną wiązanie, są znani jako monounsynowane lub monoenoi, podczas gdy ci, którzy posiadają więcej niż dwa, są znani jako polienikowe lub wielonienasycone.
Może ci służyć: polimeraza DNAObecność podwójnych wiązań jest powszechna między atomami węgla między pozycjami 9 i 10, ale nie oznacza to, że monouinsynowane kwasy tłuszczowe nie występują z nienasyceniem w innej pozycji.
W przeciwieństwie do nasyconych, nienasycone kwasy tłuszczowe są wymienione nie z końcowej grupy karboksylowej, ale zgodnie z położeniem pierwszego podwójnego wiązania C - c. Zatem są one podzielone na dwie grupy, omega-6 lub ω6 i omega-3 lub ω3.
Kwasy omega-6 mają pierwsze podwójne wiązanie w kwasach węgla 6, a kwasy omega-3 mają go w węglu 3. Nominowanie Ω jest podawane przez podwójne wiązanie najbliższe końcowej grupie metylowej.
Podwójne łącza można również znaleźć w dwóch konfiguracjach geometrycznych znanych jako "Cis " I „trans”.
Większość naturalnych nienasyconych kwasów tłuszczowych ma konfigurację „Cis”A podwójne wiązania kwasów tłuszczowych obecnych w tłuszczach komercyjnych (uwodornowanych) można znaleźć w „trans”.
W wielonienasyconych kwasach tłuszczowych dwa podwójne wiązania są zwykle oddzielone od siebie przynajmniej przez grupę metylową, to znaczy atom węgla przyłączony do dwóch atomów wodoru.
Funkcje
Kwasy tłuszczowe mają wiele funkcji w organizmach żywych i, jak wspomniano powyżej, jedna z ich istotnych funkcji jako istotnej części lipidów, które są głównymi składnikami błon biologicznych i jednej z trzech najliczniejszych biomolekuł w organizmach w organizmach żyjących w połączeniu z żywymi organizmami w połączeniu z organizmami żyjącymi z organizmami żyjącymi białka i węglowodany.
Są to również doskonałe substraty energetyczne dzięki, na które duże ilości energii są uzyskiwane w postaci ATP i innych metabolitów pośrednich.
W związku z faktem, że zwierzęta na przykład nie są w stanie przechowywać węglowodanów, kwasy tłuszczowe reprezentują główne źródło magazynowania energii pochodzącego z utleniania nadmiaru cukrów.
Krótkie kwasy tłuszczowe nasycone w okrężnicy uczestniczą w stymulacji wchłaniania wody i jonów sodu, chlorku i wodorowęglanu; Ponadto mają funkcje w produkcji śluzu, w proliferacji kolonocytów (komórki okrężnicy) itp.
Nienasycone kwasy tłuszczowe są szczególnie obfite w jadalnych olejkach pochodzenia roślinnego, które są ważne w diecie wszystkich ludzi.
 Oleje, które spożywamy codziennie, to kwasy tłuszczowe (źródło: StevePB, przez pixabay.com)
Oleje, które spożywamy codziennie, to kwasy tłuszczowe (źródło: StevePB, przez pixabay.com) Inni uczestniczą jako ligandy niektórych białek z aktywnością enzymatyczną, więc są one ważne w odniesieniu do ich wpływu na metabolizm energetyczny komórek, w którym się znajdują.
Biosynteza
Degradacja kwasów tłuszczowych jest znana jako β-utlenianie i występuje w mitochondriach komórek eukariotycznych. Przeciwnie, biosynteza występuje w cytosolu komórek zwierzęcych i w chloroplastach (fotosyntetycznych organellach) komórek roślinnych.
Jest to proces acetylo-CoA, malonylo-CoA i NADPH, występuje we wszystkich żywych organizmach i na „lepszych” zwierzętach, takich jak ssaki. Na przykład jest to bardzo ważne w tkankach wątroby i tłuszczowej, a także w gruczołach sutkowych.
NADPH zastosowany w tej trasie jest głównie produktem zależnych od NADP reakcji utleniania trasy fosforanu pentozowego, podczas gdy acetylo-CoA może pochodzić z różnych źródeł, na przykład z oksydacyjnej dekarboksylacji pirogronianu, Krebs i β- Utlenianie kwasów tłuszczowych.
Droga biosyntezy, podobnie jak β-utlenianie, jest wysoce regulowana we wszystkich komórkach przez efektory Alestheryczne i kowalencyjne modyfikacje enzymów, które uczestniczą w regulacji.
-Synteza malonylo-CoA
Trasa rozpoczyna się od tworzenia metabolicznego pośrednia.
Ta reakcja jest reakcją na dodatek do cząsteczki karboksylowej (-COH, karboksylacja) zależna od biotyny i występuje w dwóch etapach:
- Po pierwsze, przeniesienie, zależne od ATP, karboksylu pochodzącego od wodorowęglanu (HCO3-) do cząsteczki biotyny, która jest stwierdzona jako grupa protetyczna (nieproteinowa) związana z karboksylazą acetylo-CoA.
- Następnie CO2 jest przenoszony do acetylo-CoA i wytwarzany jest malonylo-CoA.
-Reakcje trasy
U zwierząt tworzenie łańcuchów węglowodanów kwasów tłuszczowych zachodzi dalej poprzez sekwencyjne reakcje kondensacyjne katalizowane przez enzym wielofrytyczny i wielofunkcyjny, znany jako syntaza kwasów tłuszczowych.
Ten enzym katalizuje kondensację jednostki acetylo-CoA i cząsteczek wielo-korek, które są wytwarzane z reakcji karboksylazy acetylo-CoA, procesu, w którym cząsteczka CO2 jest uwalniana dla każdego dodanego malonylo-CoA.
Rosnące kwasy tłuszczowe są sterygowane do białka zwanego „białkiem nośnikiem acylowym” lub ACP, które tworzy tioés z grupami acylowymi. W I. coli To białko jest polipeptydem 10 kDa, ale u zwierząt jest częścią syntazy kompleksu kwasów tłuszczowych.
Może ci służyć: noxaZerwanie tych wiązań Tioéster uwalnia duże ilości energii, co umożliwia, termodynamicznie.
Syntaza kompleksu kwasu tłuszczowego
W bakteriach aktywność kwasu syntazy faktycznie odpowiada sześciu niezależnym enzymom, które wykorzystują acetylo-CoA i malonylo-CoA do tworzenia kwasów tłuszczowych i z którymi powiązane jest sześć różnych aktywności enzymatycznej.
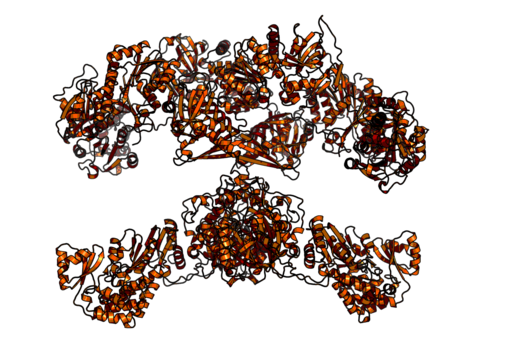 Homodimeryczna i wielofunkcyjna złożona syntaza kwasów tłuszczowych zwierząt (źródło: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons)
Homodimeryczna i wielofunkcyjna złożona syntaza kwasów tłuszczowych zwierząt (źródło: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons) Z drugiej strony u ssaków syntaza kwasu tłuszczowego jest wielofunkcyjnym homodymicznym kompleksem enzymatycznym o masie cząsteczkowej około 500 kDa, który ma sześć różnych aktywności katalitycznej i z którym związane jest białko nośnika kwasu.
Krok 1: Reakcja kroplica
Grupy tiolowe w odpadach cysteinowych odpowiedzialne za połączenie pośredników metabolicznych na enzym ACP muszą być załadowane, przed rozpoczęciem syntezy, z niezbędnymi grupami kwasowymi.
Aby to zrobić, grupa acetylowa acetylo-CoA jest przenoszona do grupy tiol (-SH) jednej z reszt cysteinowych podjednostki ACP syntazy kwasu tłuszczowego. Ta reakcja jest katalizowana przez podjednostkę ACP-acyl-transferaza.
Grupa acetylowa jest następnie przenoszona z ACP do innej reszty cysteiny w miejscu katalitycznym innej enzymatycznej podjednostki kompleksu znanego jako β-cetoacylo-acp-sinta. Zatem kompleks enzymatyczny jest „jęczmieniem”, aby rozpocząć syntezę.
Krok 2: Transfer jednostek malonylo-CoA
Malonylo-CoA, który jest wytwarzany przez acetylo-CoA Carboxilasa, jest przenoszony do grupy tiol w ACP i podczas tej reakcji część CoA jest utracona. Reakcję jest katalizowana przez podjednostkę malonil-ACP-transferazy kompleksu syntazy kwasu tłuszczowego, która następnie wytwarza malonil-ACP.
Podczas tego procesu grupa maloniczna jest powiązana z ACP i β-cotoacil-acp-sita odpowiednio za pośrednictwem łącza estrowego i innego sulphidl.
Krok 3: Kondensacja
Enzym β-zoacyl-ACP-suntaza katalizuje przeniesienie grupy acetylowej, która dołączyła do niego w przejściu „jęczmienia” w kierunku węgla 2 grupy Malonilo, która w poprzednim kroku przeniesiono do ACP.
Podczas tej reakcji uwalniana jest cząsteczka CO2, która pochodzi z malonylu, która odpowiada CO2 przyczynionego przez wodorowęglan w reakcji karboksylacji karboksylazy acetylo-CoA. Następnie występuje acetoacetylo-ACP.
Krok 4: Redukcja
Podjednostka β-zoacyl-acp-reduktazy katalizuje zmniejszenie zależnego od NADPH Aceoacetylo-ACP.
Krok 5: Odwodnienie
Ten etap tworzy trans-α, β-acyl-ACP lub ∆2-insaturado-acyl-acp (kratonil-acp), iloczyn odwodnienia d-β-hydroksybutiril-acp przez działanie gniewnego podjednostki ACP -Hydratasa.
Później kratonil-ACP jest redukowana do butiril-acp przez reakcję zależną od NADPH katalizowaną przez podjednostkę gniewnie-ac-reduktazę. Z tą reakcją pierwszą z siedmiu cykli potrzebnych do wytworzenia Palmitail-ACP, która jest prekursorem prawie wszystkich kwasów tłuszczowych.
Jak następują kolejne reakcje kondensacji?
Grupa Butirilo jest przenoszona z ACP do grupy tiol reszty cysteinowej w β-cetoacilu-acp-sita, dzięki czemu ACP jest w stanie zaakceptować inną grupę maloniczną z malonyl-CoA.
W ten sposób reakcja, która się zdarza, jest kondensacja malonil-ACP z Butturil-β-cotoacil-acp-sinta, która powoduje powstanie β-Zotohekanoil-ACP + CO2 CO2.
Palmitail-ACP, który wynika z kolejnych etapów (po dodaniu 5 kolejnych jednostek malonicznych) może być uwalniane jako wolny kwas palmitynowy dzięki aktywności enzymatycznej tioesterazy, można ją przenieść do COA lub włączyć do kwasu fosfatydowego dla dla droga syntezy fosfolipidów i triacyloglicerydów.
 Struktura kwasu palmitynowego (źródło: Andel, przez Wikimedia Commons)
Struktura kwasu palmitynowego (źródło: Andel, przez Wikimedia Commons) Syntaza kwasów tłuszczowych większości organizmów zatrzymuje się w syntezie palmitail-acp, biorąc pod uwagę, że miejsce katalityczne podjednostki β-cetoacylo-acp-syntazy ma konfigurację, w której można uwzględnić tylko kwasy tłuszczowe o tej długości.
W jaki sposób kwasy tłuszczowe o nieparzystych atomach węgla?
Są one stosunkowo powszechne w organizmach morskich i są również syntetyzowane przez złożoną syntazę kwasów tłuszczowych. Jednak reakcja „jęczmienia” zachodzi z dłuższą cząsteczką, propionil-ACP, z trzech atomów węgla.
Gdzie i jak mają kwasy tłuszczowe dłuższych łańcuchów?
Kwas palmitynowy, jak skomentowano, służy jako prekursor wielu nasyconych i nienasyconych kwasów tłuszczowych o dłuższych łańcuchach. Proces „wydłużenia” kwasów tłuszczowych występuje w mitochondriach, podczas gdy wprowadzenie nienasycenia występuje zasadniczo w retikulum endoplazmatycznym.
Wiele organizmów zamienia ich nasycone kwasy tłuszczowe w nienasycone jako adaptacja do niskich temperatur środowiskowych, ponieważ pozwala im to utrzymać punkt fuzji lipidów poniżej temperatury otoczenia.
Właściwości kwasu tłuszczowego
Wiele właściwości kwasów tłuszczowych zależy od ich długości łańcucha oraz obecności i liczby nienasycenia:
Może ci służyć: abiotyczna teoria syntezy: główne cechy- Nienaturowane kwasy tłuszczowe mają niższe temperatury topnienia niż nasycone kwasy tłuszczowe o tej samej długości.
- Długość kwasów tłuszczowych (liczba atomów węgla) jest odwrotnie proporcjonalna do płynności lub elastyczności cząsteczki, to znaczy, że najbardziej „krótkie” cząsteczki są bardziej płynne i odwrotnie.
Ogólnie rzecz biorąc, płynne substancje tłuszczowe składają się z kwasów tłuszczowych o krótkich łańcuchach i obecności nienaturalnej.
Rośliny mają obfitą ilość nienasyconych kwasów tłuszczowych, a także zwierzęta żyjące w bardzo niskich temperaturach, ponieważ te, jako składniki lipidów obecnych w błonach komórkowych, dają im większą płynność w tych warunkach.
W warunkach fizjologicznych obecność podwójnego wiązania w łańcuchu węglowodorowym tłuszczu.
Obecność podwójnych wiązań w kwasach tłuszczowych związanych z cząsteczkami lipidów ma bezpośredni wpływ na stopień „opakowania”, jaki mogą mieć w błonach, do których należą, a wraz z nimi mają również wpływ na białka błonowe.
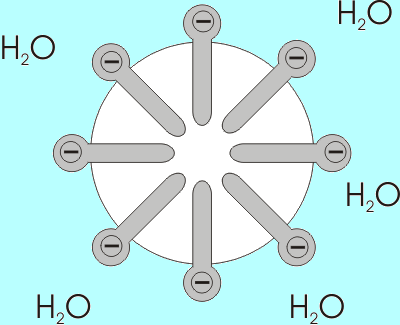 Przykład tworzenia Myli kwasów tłuszczowych z grupami karboksylowymi narażonymi na środowisko wodne (źródło: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licencje/by-sa/3.0/)] przez Wikimedia Commons)
Przykład tworzenia Myli kwasów tłuszczowych z grupami karboksylowymi narażonymi na środowisko wodne (źródło: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licencje/by-sa/3.0/)] przez Wikimedia Commons) Rozpuszczalność kwasów tłuszczowych maleje wraz ze wzrostem długości łańcucha, więc są one odwrotnie proporcjonalne. W mieszaninach wodnych i lipidów kwasy tłuszczowe są powiązane w strukturach znanych jako micele.
Micela to struktura, w której alifatyczne łańcuchy kwasów tłuszczowych są „zamknięte”, a tym samym „wydalanie” wszystkich cząsteczek wody i na której powierzchni są grupy karboksyl.
Nomenklatura
Nomenklatura kwasów tłuszczowych może być nieco złożona, zwłaszcza jeśli odnosi się do zwykłych nazwisk, które otrzymują, które często są powiązane z pewną właściwością fizykochemiczną, z miejscem, w którym są lub inne cechy cechy.
Wielu autorów uważa, że jako dzięki końcowej grupie karboksylowej cząsteczki te są jonizowane do fizjologicznego pH, należy je nazywać „karboksylami” za pomocą zakończenia ”Ato ".
Według systemu IUPAC wyliczenie atomów węgla tłuszczowego. Łańcuch zaciskowy metyl zawiera atom węgla ω.
Ogólnie rzecz biorąc, w systematycznej nomenklaturze otrzymują nazwę „rodzicielskiego” węglowodorów (węglowodór z tą samą liczbą atomów węgla), a ich zakończenie jest zastąpione ”albo" przez „OICO”, Jeśli jest to nienasycony kwas tłuszczowy, dodaje się zakończenie "Mgła".
Rozważmy na przykład przypadek kwasu tłuszczowego 18 atomów węgla (C18):
- Ponieważ węglowodór z taką samą ilością atomów węgla jest znany jako oktadecano, nazywany jest kwas nasycony ”Kwas oktadekanowy„O, dobrze”Octadecanoato”A jego zwykłą nazwą to kwas stearynowy.
- Jeśli masz podwójne wiązanie między niektórymi parami atomów węgla w jego strukturze, jest on znany jako "Kwas oktadecenowy"
- Jeśli masz dwa podwójne res c - c, to jest nazywane „Kwas oktadecadienoinowy” A jeśli masz trzy "Kwas oktadecatrienoinowy".
Jeśli chcesz podsumować nomenklaturę, użyj 18: 0 dla 18 -węglowych kwasów tłuszczowych i braku podwójnego wiązania (nasycone) i, w zależności od stopnia nienasycenia, zamiast zero zapisuje się 18: 1 dla cząsteczki z cząsteczką z nienaturowaniem, 18: 2 dla jednego z dwiema nienaturowaniem i tak dalej.
Jeśli chcesz określić między tym, które atomy węgla są podwójnymi wiązaniami w nienasyconych kwasach tłuszczowych, symbol ∆ jest używany z badaniem numerycznym, które wskazuje na miejsce nienasycenia i prefiks „Cis" albo „trans”, w zależności od konfiguracji tego.
Bibliografia
- Badui, s. (2006). Chemia gastronomiczna. (I. Quintanar, wyd.) (Czwarty ed.). Meksyk d.F.: Edukacja Pearsona.
- Garrett, r., I Grisham, C. (2010). Biochemia (Wydanie 4.). Boston, USA: Brooks/Cole. Cengage Learning.
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biochemia (3 wyd.). San Francisco, Kalifornia: Pearson.
- Murray, r., Bender, d., Botham, k., Kennelly, s. 1., Rodwell, v., I Weil, p. (2009). Ilustrowana biochemia Harpera (28. wyd.). McGraw-Hill Medical.
- Nelson, zm. L., & Cox, m. M. (2009). Zasady biochemii lehninger. Omega Editions (Ed.).
- Rawn, J. D. (1998). Biochemia. Burlington, Massachusetts: Neil Patterson Publishers.
- TVZICKA, e., Kremmyda, L., Stankova, ur., & Zak, do. (2011). Kwasy tłuszczowe jako biokomponcje: ich rola w metabolizmie ludzkim, zdrowiu i chorobie- przegląd. Część 1: Klasyfikacja, źródła diety i funkcje biologiczne. Biomed Pap Med Fare Univ Palacky Olomouc Czech Repub, 155(2), 117-130.
- « Struktura, funkcje, klasy i przykłady glikoprotein
- Wpływ przynależności do obywatelstwa cyfrowego »