

Struktura kardiolipiny, synteza, funkcje
- 4678
- 668
- Estera Wojtkowiak
Kardiolipina, Znany również jako defatydyloglicerol, jest lipidem rodziny gliceofosfolipidów i grupy poliglifosfolipidów. Występuje w błonie mitochondrialnej organizmów eukariotycznych, w błonie plazmatycznej wielu bakterii, a także w niektórych łukach.
Został odkryty przez Pangborn w 1942 r. Na podstawie analizy lipidów błony tkanki serca bydła. Jego struktura została zaproponowana w 1956 roku, a synteza chemiczna miała miejsce około 10 lat później.
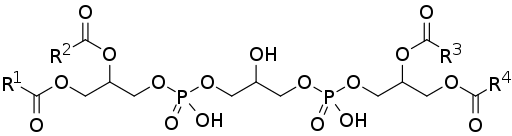
Struktura kardiolipiny (źródło: Edgar181 [domena publiczna] za pośrednictwem Wikimedia Commons) Niektórzy autorzy uważają, że ich obecność jest ograniczona do błon wytwarzających ATP, podobnie jak w przypadku mitochondriów u eukariotów, błon plazmatycznych u bakterii i wodorosomów (organelli typu Mmitokondriów) w niektórych protestacjach.
Fakt, że kardiolipina znajduje się w mitochondriach i w błonie bakterii w osoczu, zastosowano do wzmocnienia fundamentów teorii endosimbiotycznej, która stała się zależna od mitochondrii eukariotów przez fagocytozę bakterii, która następnie stała się zależna komórka i odwrotnie.
Jego biosyntetyczna droga u zwierząt została opisana w latach 1970–1972, a następnie wykazano, że jest to ta sama droga, która występuje w roślinach, drożdżach, grzybach i bezkręgowcach. Nie jest to bardzo obfity lipid, ale komórki wymagają, aby funkcjonować prawidłowo.
Znaczenie tego fosfolipidów dla mitochondriów, a zatem dla metabolizmu komórkowego, jest widoczne, gdy wadliwe funkcjonowanie dróg metabolicznych związanych z nim powoduje ludzką patologię znaną jako zespół Bartha (miopatia kardiooskkieletowa).
[TOC]
Struktura
Kardiolipina lub defatezatydyloglicerolu składa się z dwóch kwasów fosfatydowych (łatwiejszych fosfolipidów) połączonych ze sobą przez cząsteczkę glicerolu.
Kwas fosfatydowy, jeden z wspólnych pośredników na biosyntetycznych drogach innych fosfolipidów, składa się z 3-fosforanowej cząsteczki glicerolu, do której dwa łańcuchy kwasów tłuszczowych są esteryfikowane w pozycjach węglowodanów 1 i 2, więc jest ona również znana jako 1,2 -Diacyloglicerol 3-fosforan.
Może ci służyć: wtórna struktura białek: charakterystykaDlatego kardiolipina składa się z trzech cząsteczek glicerolu: centralnego glicerolu, wraz z grupą fosforanową w węglu 1, kolejnej grupie fosforanowej w węglu 3 i grupie hydroksylowej w węglu 2; i dwie „boczne” glicerenie.
Dwie „boczne” cząsteczki glicerolu wiążą się z cząsteczką centralną przez „mosty glicerolu” przez ich węgle w pozycji 3. W węglach pozycji 1 i 2, dwa łańcuchy kwasów tłuszczowych i zmienne nasycenie estryfikowały.
Kardiolipina jest lipidem, który może, ale nie musi tworzyć bicapas, w zależności od obecności lub braku kationów. Jest to związane z faktem, że jest to symetryczna cząsteczka, co czyni ją ważną w błonach odpowiedzialnych za procesy przekazywania energii.
Podobnie jak inne lipidy grupy poliglifosfolipidów, kardiolipina ma kilka grup hydroksylowych, które mogą służyć dla połączenia kwasów tłuszczowych. Dlatego ma wiele pozycyjnych stereoizomerów.
Jego kwasy tłuszczowe
Różne badania wykazały, że kwasy tłuszczowe przyłączone do bocznych cząsteczek glicerolu kardiolipiny są zwykle nienasycone, ale stopień nienasycenia nie został określony.
Następnie takie kwasy tłuszczowe mogą mieć od 14 do 22 węgli o długości i od 0 do 6 podwójnych wiązań. To i fakt, że kardiolipina ma cztery powiązane cząsteczki tłuszczu, oznacza, że może istnieć wiele zmiennych i kombinacji tego fosfolipidu.
Synteza
Biosynteza kardiolipiny, jak można się było spodziewać, zaczyna się od syntezy kwasu fosfatydowego lub 3-fosforanu 1,2-dicyloglicerolu z 3-fosforanu i kwasów tłuszczowych i kwasów tłuszczowych i kwasów tłuszczowych. Proces ten występuje w mitochondriach eukariotów i w błonie plazmatycznej bakterii.
Może ci służyć: Cyclasa Adenilate: Charakterystyka, typy, funkcjeSynteza u eukariotów
Po utworzeniu kwas fosfatydowy reaguje z cząsteczką o wysokiej energii analogicznej do ATP: CTP. Następnie pośrednik, również o wysokiej energii, jest znany jako fosfatydylo-CMP. Aktywowana grupa fosfatydilowa jest przenoszona do grupy hydroksylowej w pozycji C1 centralnej cząsteczki glicerolu, która służy jako szkielet.
Ten proces wynika. Otrzymuje to kolejną grupę fosfatydylową aktywowaną z innej cząsteczki fosfatydyl-CMP, reakcję katalizowaną przez fosfatydylotransferazę znaną również jako syntaza kardiolipiny.
Syntaza enzym kardiolipiny leży w wewnętrznej błonie mitochondrialnej i wydaje się tworzyć wielki kompleks, przynajmniej w drożdżach. Jego gen jest wyrażany w dużych ilościach w bogatych tkaninach w mitochondriach, takich jak serce, wątroba i mięsień szkieletowy kręgowców.
Regulacja jego aktywności zależy w dużej mierze od tych samych czynników transkrypcyjnych i czynników hormonalnych, które modulują biogenezę mitochondriów.
Po zsyntetyzowaniu w wewnętrznej błonie mitochondrialnej, kardiolipina musi zostać przeniesiona w kierunku zewnętrznej membrany mitochondrialnej, aby podano serię procesów topologicznych w błonie, a inne tego same elementy strukturalne są zakwaterowane.
Synteza u prokariotów
Zawartość kardiolipiny w bakteriach może być bardzo zmienna i zależy głównie od stanu fizjologicznego komórek: zwykle jest mniej obfita w wykładniczej fazie wzrostu i bardziej obfita, gdy nastąpi jego zmniejszenie (na przykład w fazie stacjonarnej).
Jego biosyntetyczna trasa może być nakręcona przez różne stresujące bodźce, takie jak deficyt energii lub stres osmotyczny.
Aż do tworzenia fosfatydyloglicerolu, proces eukariotów i prokariotów jest równoważny, ale w prokariotach fosfatydyloglicerolu, przez transcensizację, grupę fosfatydylową z innej cząsteczki fosfatydyloglicerolu. Ta reakcja jest katalizowana przez enzym fosfolipazę D znaną również jako syntaza kardiolipiny.
Może ci służyć: SGLT (białka transportu glukozy sodu)Ta reakcja jest znana jako reakcja „transfatidilation” (z angielskiego ”Transposfatydylacja ”), Gdzie jedna z fosfatydyloglicereno działa jako dawca grupy fosfatydilowej, a drugi jako akceptor.
Funkcje
Fizyczne cechy cząsteczek kardiolipiny najwyraźniej pozwalają na niektóre interakcje, które pełnią ważne funkcje w organizacji strukturalnej błon, w których się znajdują.
Wśród tych funkcji jest dyskryminacja niektórych domen błonowych, interakcja lub „krzyżowanie” z białkami transmarketowymi lub ich subdomenami, między innymi.
Dzięki jego charakterystyce fizykochemicznej kardiolipina jest rozpoznawana jako lipid, który nie tworzy Bápara, ale którego funkcją może być stabilizacja i „pomieścić” białka transbłonowe w lipidowych dwuosobowych dwuosobowych.
Szczególnie jego charakterystyka elektryczna daje funkcje w procesach transferu protonów, które zachodzą w mitochondriach.
Chociaż komórki mogą przetrwać bez tego fosfolipidu, niektóre badania wykazały, że jest to wymagane do ich optymalnego funkcjonowania.
Bibliografia
- Harayama, t., & Riezman, H. (2018). Zrozumienie różnorodności składu lipidów błonowych. Nature przegląd biologii komórek molekularnych, 19(5), 281-296.
- Luckey, m. (2008). BIOLOGIA Membrana strukturalna: z podstawami biochemicznymi i biofizycznymi. Cambridge University Press.
- Murray, r., Bender, d., Botham, k., Kennelly, s. 1., Rodwell, v., I Weil, p. (2009). Ilustrowana biochemia Harpera (28. wyd.). McGraw-Hill Medical.
- Van Meer, G., Voelker, zm. R., & Feigenson, G. W. (2008). Lipidy membrane: gdzie są i jak się zachowują. Recenzje przyrody, 9, 112-124.
- Vance, J. I., & Vance, D. I. (2008). Biochemia lipidów, lipoprotein i błon. W Nowe zrozumienie Biochemistry vol. 36 (Wydanie 4.). Elsevier.