

Trzy cechy i funkcje w organizmie
- 821
- 173
- Marianna Czarnecki
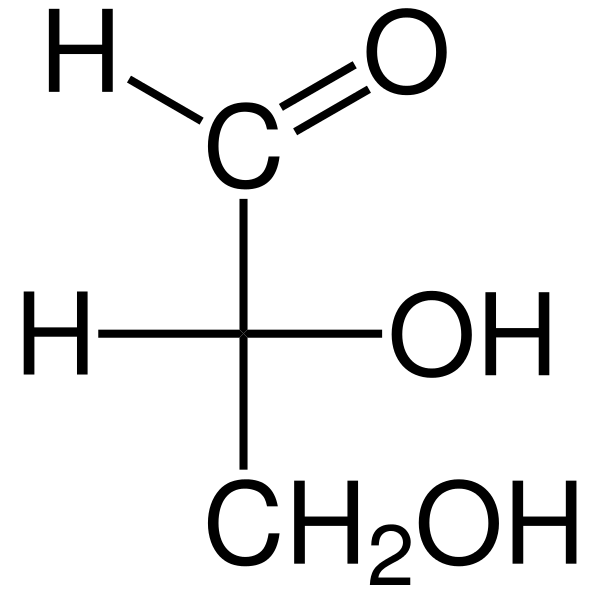
Triozie Są to trzy -węglowe monosacharydy, których empiryczna wzór chemiczny to C3H6ALBO6. Istnieją dwa trio: gliceraldehyd. Triozy są ważne w metabolizmie, ponieważ łączą trzy szlaki metaboliczne: glikolizę, glukoneogenezę i ścieżkę fosforanu pentozowego.
Podczas fotosyntezy cykl Calvin jest źródłem triozów, które służą do biosyntezy fruktozy-6-fosforan. Ten cukier w sposób fosforylowany jest przekształcany przez enzymatycznie katalizowane kroki w rezerwowe lub strukturalne polisacharydy.
Źródło: Wesalius [domena publiczna] Triozy uczestniczą w biosyntezy lipidów, które są częścią błon komórkowych i adipocytów.
[TOC]
Charakterystyka
Gliceraldehyd Aldosa ma quiralny atom węgla, a zatem ma dwa enantiomeros, L-gliceraldehyd i D-gliceraldehyd. Oba enancjomery D i L mają różne cechy chemiczne i fizyczne.
D-gliceraldehyd złamał płaszczyznę spolaryzowanego światła po prawej (+) i ma obrót [α] D, w 25 ° C, od +8.7 °, podczas gdy L-gliceraldehyd złamał płaszczyznę spolaryzowanego światła po lewej (-) i ma obrót [α] D, w 25 ° C, od -8.7 °.
Chiralny węgiel gliceraldehydu to węgiel 2 (C-2), który jest alkoholem wtórnym. Projekcja Fischera reprezentuje grupę hydroksylową (-OH) d-gliceraldehydu po prawej i grupie OH-lewej L-glyce.
Dihydroksyaceton nie ma chiralnych węgli i nie ma postaci enancjomerycznych. Dodanie grupy hydroksymetylenu (-choh) do gliceraldehydu lub dihydroksyacetonu umożliwia utworzenie nowego centrum chiralnego. W związku z tym cukier jest tetrozą, ponieważ ma cztery węgle.
Dodanie grupy -Chah do Terosy tworzy nowe centrum chiralne. Utworzony cukier to pentoza. Grupy -Chah mogą być nadal dodawane do maksymalnie dziesięciu węgli.
Może ci służyć: filogenezaFunkcje w organizmie
Triosas jako pośredników w glikolizy, glukoneogenezie i ścieżce fosforanowej pentosa
Glikoliza składa się z pęknięcia cząsteczki glukozy w dwóch cząsteczkach pirogronianu w celu wytworzenia energii. Ta droga implikuje dwie fazy: 1) faza przygotowawcza lub zużycie energii; 2) Faza wytwarzania energii. Pierwszy to ten, który produkuje trio.
W pierwszej fazie zwiększona zawartość energii bez glukozy, poprzez tworzenie fosfoésterów. W tej fazie triffosforan adenozyny (ATP) jest dawcą fosforanu. Ta faza kończy się konwersją fosfoéster fosfoasteru fosfosteru fruktozy (F1.6BP) w dwóch fosforanach triosas, fosforanu gliceraldehydu (GA3P) i dihydroksyaceton-fosforan (DHAP).
Glukoneogeneza to biosynteza glukozy z pirogronianu i innych pośredników. Użyj wszystkich enzymów glikolizy, które katalizują reakcje, których biochemiczna standardowa zmiana energii jest w równowadze (GGº '~ 0). Z tego powodu glikoliza i glukoneogeneza mają wspólne pośredniki, w tym GA3P i DHAP.
Ścieżka fosforanu pentozowego składa się z dwóch etapów: fazy oksydacyjnej glukozy-6-fosforanu i kolejnego tworzenia NADPH i rybozy-5-fosforanu. W drugiej fazie ryboza 5-fosforanowa jest przekształcana w pośredniki glikolizy, F1.6BP i GA3P.
Triozę i cykl Calvin
Fotosynteza jest podzielona na dwa etapy. W pierwszym występują reakcje zależne od światła, które wytwarzają NADPH i ATP. Substancje te są używane w drugim, w którym istnieje utrwalanie dwutlenku węgla i tworzenia się heksozy z trioów przez ścieżkę znaną jako Calvin Cycle.
W cyklu Calvina enzym rybulozy o długości 1,5 bifsygenazy (rubisco) katalizuje kowalencyjny związek Co2 Do pentozowego rybulosa 1,5-bifosforanu i łamie niestabilne pośrednie z sześciu atomów węgla w dwóch cząsteczkach trzech atomów węgla: 3-fosfoglicenianu.
Może ci służyć: oligosacharydy: cechy, skład, funkcje, typyPoprzez reakcje enzymatyczne, które obejmują fosforylację i redukcję 3-fosfoglikanu, przy użyciu ATP i NADP, GA3P występuje. Ten metabolit przekształca się w fruktozę 1,6-bifosforan (F1.6BP) przez szlak metaboliczny podobny do glukoneogenezy.
Przez działanie fosfatazy F1.6BP jest przekształcane w fruktozę-6-fosforan. Następnie fosfhhexoza izomeazy wytwarza glukozę 6-fosforan (GLC6P). Wreszcie epicheraza przekształca GLC6P w 1-fosforan glukozy, który służy do biosyntezy skrobiowej.
Trio i lipidy błon biologicznych i adipocytów
GA3P i DHAP mogą tworzyć fosforan glicerolu, który jest niezbędnym metabolitem do biosyntezy triacylogliceroli i glicerolipidów. Wynika to z faktu, że oba fosforan triosas może być połączony przez reakcję katalizowaną przez fosforan izomerazy triosa, który utrzymuje obie trio w równowadze.
Dehydrogenaza enzymu glicerolu-fosforanowego katalizuje reakcję ograniczania utleniania, w której NADH przekazuje kilka elektronów DHAP w celu utworzenia 3-fosforanowego glicerolu i NAD+. 3-fosforan L-glicerol jest częścią szkieletu fosfolipidów, które są strukturalną częścią błon biologicznych.
Glicerol jest proquiralny, brakuje mu asymetrycznych węgli, ale gdy jeden z dwóch głównych alkoholi tworzy Fosfoester, można go poprawnie nazwać 3-fosforanem L lub 3-fosforanu D lub glicerolu D lub D-glicerolu.
Glyceofosfolipidy są również nazywane fosfoglicerydami, nazywane kwasem fosfatydowym pochodnym. Fosfoglicerydy mogą tworzyć fosfoacyloglicereno poprzez utworzenie wiązań estrów z dwoma kwasami tłuszczowymi. W takim przypadku powstałym produktem jest 1,2-fosfodiacyloglicerol, który jest ważnym składnikiem błon.
Gliceofosfaza katalizuje hydrolizę 3-fosforanowej grupy fosforanowej glicerolu, wytwarzając glicerol więcej fosforanu. Glicerol może służyć jako początkowy metabolit biosyntezy triacyloglicerydów, które są powszechne w adipocytach.
Może ci służyć: test koagulazy: fundament, procedura i zastosowaniaTriozy i błony archeobakterii
Podobnie jak eubakterie i eukariotów, 3-fosforanowy glicerol powstaje z fosforanu triosas (GA3P i DHAP). Istnieją jednak różnice: pierwsza jest to, że 3-fosforan glicerolu w błonach archeobakteryjnych to L-konfiguracja, podczas gdy w błonach eubakteryjnych i eukariotycznych jest konfiguracją d.
Druga różnica polega na tym, że błony archeobakteryjne tworzą wiązania estra z dwoma długimi łańcuchami węglowodorowymi grup izoprenoidalnych, podczas gdy w eubakterie i eukariotach glicerolu tworzy wiązania estra (1,2-dialiflicerolu) z dwoma łańcuchami węglowodorowymi kwasów tłuszczowych.
Trzecia różnica polega na tym, że w błonach archeobakteryjnych zastępcze w fosforan i glicerolu 3-fosforanowym różnią się od eukarotów i eukariontów. Na przykład grupa fosforanowa jest powiązana z disacharydem α-Glucopiranso- (1®2)-β-Galatofuranosa.
Bibliografia
- Cui, s. W. 2005. Węglowodany żywnościowe: chemia, właściwości fizyczne i zastosowania. CRC Press, Boca Raton.
- Kogut, s., Mäkinen, K, Honkala i., Saag, m., Kennepohl, e., Eapen, a. 2016. Erytritol jest bardziej skuteczny niż ksylitol i sorbitol w zarządzaniu punktami końcowymi zdrowia jamy ustnej. International Journal of Dentistry.
- Nelson, zm. L., Cox, m. M. 2017. Zasady biochemii lehninger. W. H. Freeman, Nowy Jork.
- Sinnott, m. L. 2007. Chemia węglowodanów i struktura i mechanizm biochemii. Royal Society of Chemistry, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Węglowodany: niezbędne cząsteczki życia. Elsevier, Amsterdam.
- Voet, d., Voet, J. G., Pratt, c. W. 2008. Podstawy biochemii - życie na poziomie molekularnym. Wiley, Hoboken.
- « Charakterystyka aldozy, typy, liczba węgli
- Mechanizm działania i skutki uboczne fluoksetyny (Prozac) »