

Tilacoid

- 2230
- 153
- Maksymilian Kępa
Tilacoid w chloroplastu Co to są tilacoids?
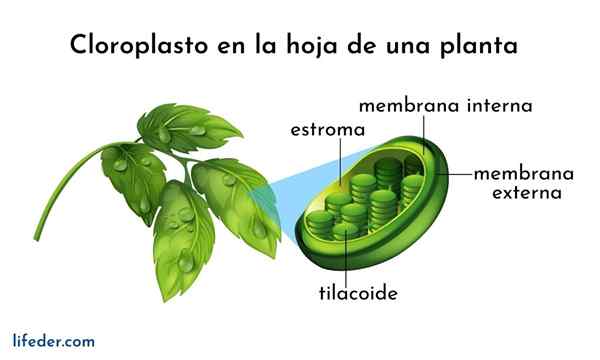
Tilacoid Są to przedziały w postaci płaskich worków znajdujących się w chloroplastach w komórkach roślinnych, w cyjanobakterii i algach. Zazwyczaj są one organizowane w strukturze zwanej Grana -Plural Granum- i wyglądaj jak stos monet.
Tilacoidy są uważane za trzeci system błony chloroplastowej, oprócz wewnętrznej i zewnętrznej błony wspomnianej organelli. Membrana tej struktury oddziela się wewnątrz tilacoidów zrębu chloroplastu i ma serię pigmentów i białek zaangażowanych w szlaki metaboliczne.
W tilacoidach istnieją niezbędne reakcje biochemiczne na fotosyntezę, proces, w którym rośliny przyjmują światło słoneczne i przekształcają je w węglowodany. W szczególności mają niezbędne maszyny zakotwiczone w błonie do wykonania fazy zależnej od światła słonecznego, w której światło jest uwięzione i przekształcane w energię (ATP) i NADPH.
Charakterystyka tilacoidów
- Tilacoidy są wewnętrznym trójwymiarowym układem błoniastym chloroplastów. W pełni dojrzałe chloroplasty mają stosy od 40 do 60 grany, o średnicy między 0,3 a 0,6 µm.
- Liczba tilacoidów, które składają się na granaty, różni się znacznie: od mniej niż 10 torebek w roślinach narażonych na wystarczającą ilość światła słonecznego, do ponad 100 tilacoidów w roślinach żyjących w środowiskach o ekstremalnym cieniu.
- Ułożone tilacoidy są ze sobą połączone, tworząc ciągły przedział w chloroplast. Wnętrze Tilacoid jest dość przestronnym przedziałem natury wodnej.
- Membrana Tilacoid jest niezbędna do fotosyntezy, ponieważ odbywa się tam pierwszy etap procesu.
Struktura Tilacoid
Tilacoids to struktury dominujące w całkowicie dojrzałym chloroplastie. Jeśli chloroplast jest wyświetlany w tradycyjnym mikroskopie optycznym, można zaobserwować gatunki ziaren.
Są to stosowanie tilacoidów; Dlatego pierwsi obserwatorzy tych struktur nazywano „grana”.
Za pomocą mikroskopu elektronicznego obraz mógł zostać rozszerzony i stwierdzono, że natura tych ziaren była naprawdę ułożona w stos.
Powstawanie i struktura błony tilacoidowej zależy od tworzenia chloroplast. Obecność światła stymuluje konwersję do chloroplastów, a następnie tworzenie ułożonych tilacoidów.
Może ci służyć: nasienie: funkcje, części, cykl życiaMembrana Tilacoid
W chloroplastach i cyjanobakterii błona tilacoidalna nie kontaktuje się z wewnętrzną częścią błony plazmatycznej. Jednak tworzenie błony tilacoidowej zaczyna się od inwazji wewnętrznej błony.
W cyjanobakterii i niektórych gatunkach glonów tilacoidy są tworzone przez pojedynczą warstwę lamelli. Natomiast istnieje bardziej złożony system w dojrzałych chloroplastach.
W tej ostatniej grupie można rozróżnić dwie podstawowe części: Grana i Lamella Stroma. Pierwszy składa się z małych albumów, a drugi jest odpowiedzialny za połączenie tych stosów ze sobą, tworzenie konstrukcji trwa: światło tilacoid.
Skład lipidowy błony
Lipidy tworzące membranę są wysoce wyspecjalizowane i składają się z prawie 80 % diacyloglicerolu galaktozylu: monogalaktozyloglicerolu i digiloglicerolu diglaktozylowego. Te galaktolipidy mają bardzo nienasycone łańcuchy, typowe dla tilacoidów.
Podobnie błona tilacoidalna zawiera lipidy, takie jak fosfatydyloglicerolu w mniejszej proporcji. Wspomniane lipidy nie są rozmieszczone jednorodnie w obu warstwach błony; Istnieje pewien stopień asymetrii, który wydaje się przyczyniać się do działania struktury.
Skład białka błony
Fotosystemy I i II są dominującymi składnikami białkowymi w tej błonie. Są one związane z kompleksem cytochromu B6Syntaza F i ATP.
Stwierdzono, że większość elementów Photosystem II znajduje się w zapakowanych błonach Grana, podczas gdy fotosystem I znajduje się głównie w unantiled Tilacoid Membranes. Oznacza to, że istnieje fizyczna separacja między obiema fotosystemami.
Kompleksy te obejmują kompleksowe białka błonowe, białka obwodowe, kofaktory i różne pigmenty.
Tilacoid Pirmes
Wnętrze tilacoidów składa się z substancji wodnej i grubej, której skład różni się od zrębu. Uczestniczyć w fotofosforylacji, przechowując protony, które wygenerują siłę proton-motor do syntezy ATP. W tym procesie pH Lumen może osiągnąć 4.
W proteoma Lummen modelu organizmu Thalian Arabidopsis Zidentyfikowano ponad 80 białek, ale ich funkcje nie zostały całkowicie wyjaśnione.
Białka światła biorą udział w regulacji biogenezy tilacoidowej oraz w aktywności i zastąpieniu białek, które tworzą kompleksy fotosyntetyczne, zwłaszcza fotosystem II i NAD (P) H d edrogensa.
Może ci służyć: Promielocito: Charakterystyka, funkcje i patologieFunkcje tilacoid
Proces fotosyntezy, niezbędny dla warzyw, zaczyna się w tilacoidach. Membrana, która ich wyznacza zrębem chloroplastów.
Etapy fotosyntezy
Fotosyntezę można podzielić na dwa wielkie stadia: reakcje lekkie i ciemne reakcje.
Jak sama nazwa wskazuje, reakcje należące do pierwszej grupy mogą przebiegać tylko w obecności światła, podczas gdy reakcje drugiej grupy mogą pojawić się z tym. Zauważ, że środowisko nie jest „ciemne”, jest ono niezależne od światła.
Pierwsza grupa reakcji, „światło”, występuje w tilacoidie i można ją podsumować w następujący sposób: światło + chlorofilu + 12 godzin2O + 12 NADP+ + 18 ADP + 18 PSiema à 6 o2 + 12 NADPH + 18 ATP.
Druga grupa reakcji występuje w zrębie chloroplastu i przyjmuje ATP i NADPH zsyntetyzowane w pierwszym etapie w celu zmniejszenia węgla dwutlenku węgla do glukozy (C6H12ALBO6). Drugi etap można podsumować w: 12 NADPH + 18 ATP + 6 CO2 à c6H12ALBO6 + 12 NADP+ + 18 ADP + 18 PSiema + 6 godz2ALBO.
Zależny od światła etap
Reakcje świetlne obejmują szereg struktur zwanych fotosystemami, które znajdują się w błonie tilacoidowej i zawierają wewnątrz 300 cząsteczek pigmentowych, wśród tych chlorofilu.
Istnieją dwa rodzaje fotosystemu: pierwszy ma maksymalny pik absorpcji światła wynoszący 700 nanometrów i jest znany jako P700, Podczas gdy drugi nazywa się P680. Oba są zintegrowane z błoną tilacoidalną.
Proces rozpoczyna się, gdy jeden z pigmentów pochłania foton i ten „podskakiwanie” wobec innych pigmentów. Gdy cząsteczka chlorofilu pochłania światło, elektron przeskakuje, a inna cząsteczka go pochłania. Cząsteczka, która straciła elektron, jest teraz utleniona i ma obciążenie ujemne.
P680 Złap energię światła z chlorofilu a. W tym fotosystemie elektron jest wrzucony do bardziej energetycznego systemu do pierwotnego akceptora elektronów.
Ten elektron spada na fotosystem I, przez łańcuch przenośnika elektronów. Ten system reakcji utleniania i realizacji jest odpowiedzialny za przenoszenie protonów i elektronów z jednej cząsteczki do drugiej.
Może ci służyć: limfoblastyInnymi słowy, istnieje przepływ elektronów od wody do fotosystemu II, fotosystemu I i NADPH.
Fotofosforylacja
Część protonów generowanych przez ten układ reakcji znajduje się wewnątrz tilacoid (zwany także Tilacoid LUZ), tworząc gradient chemiczny, który generuje siłę protonową.
Protony przenoszą się z przestrzeni Tilacoid do zrębu, korzystnie po gradiencie elektrochemicznym; to znaczy opuszczają tilacoid.
Jednak przejście protonów nie jest nigdzie w błonie, muszą to zrobić poprzez złożony układ enzymatyczny zwany syntezazą ATP.
Ten ruch protonu w kierunku zrębu. Synteza ATP za pomocą światła nazywa się fotofosforylacją.
Te wspomniane etapy występują jednocześnie: chlorofil Photosystem II traci elektron i musi zastąpić go elektronem z pęknięcia cząsteczki wody; Fotosystem, który łapie światło, utlenia i uruchamia elektron złapany przez NADP+.
Zaginionym elektronem fotosystemu I zostaje zastąpiony przez wynik fotosystemu II. Związki te zostaną zastosowane w kolejnych reakcjach utrwalania węgla, w cyklu Calvin.
Ewolucja
Ewolucja fotosyntezy jako procesu liberowania tlenu pozwoliła na życie, jakie znamy.
Proponuje się, aby fotosynteza została opracowana kilka miliardów lat temu w przodku, który dał początek obecnej sinice, w oparciu o skromny kompleks fotosyntetyczny.
Proponuje się, aby ewolucji fotosyntezy towarzyszyły dwa niezbędne wydarzenia: tworzenie fotosystemu p680 oraz geneza wewnętrznego układu błony, bez połączenia z błoną komórkową.
Istnieje niezbędne białko VIPP1 do tworzenia tilacoides. Rzeczywiście, to białko jest obecne w roślinach, algach i sinice, ale nieobecne w bakteriach, które wykonują anoksyczną fotosyntezę.
Uważa się, że ten gen może pochodzić z powielania genów w możliwym przodku Cyanobakterii. Istnieje tylko jeden przypadek cyjanobakteryjny, który jest w stanie wykonywać fotosyntezę tlenu i nie ma tilacoides: gatunek Gloeobacter voolaceus.