

Główne teorie dziedzictwa postmendelskiego

- 3562
- 192
- Eliasz Dubiel
Postmendelian Heritage Obejmuje wszystkie pomysły i/lub dzieła, które powstały po badaniach opublikowanych przez religijnego przyrodnika Gregorio Mendla w 1865 r., A które wyjaśniają zjawiska lub zachowania genetyczne różniące się od tych opisanych przez to.
Gregorio Mendel jest uważany za ojca genetyki, ponieważ badanie krzyżów między roślinami grochu ustanowiło pierwsze prawa dziedzictwa ponad 100 lat temu. Pierwsze prawo, zwane „prawem segregacji”, sugeruje, że geny mogą istnieć w różnych lub alternatywnych formach.
 Gregorio Mendel (Źródło: Nieznany autor / domena publiczna, Via Wikimedia Commons)
Gregorio Mendel (Źródło: Nieznany autor / domena publiczna, Via Wikimedia Commons) Te alternatywne formy nazywane są „allelami” i bezpośrednio wpływają na fenotypowe postacie gatunku. Allele mogą określić kolor włosów, oczy, rodzaj włosów itp.
W swoim pierwszym prawie Mendel zaproponował, aby każda cecha fenotypowa przekazywana od rodziców na ich dzieci jest kontrolowana przez dwa allele, każdy od rodzica (jeden z „matki” i drugi „ojca”).
Zgodnie z tym, gdy rodzicielstwo zapewniają różne allele, potomstwo powstaje w mieszaninie, w której dominuje jeden z alleli (ten, który jest wyrażany w fenotypie), a drugi jest recesywny (nie jest widoczny lub jest „utajone ”).
Drugie prawo Mendla jest znane jako „Prawo niezależnego dziedziczenia”, ponieważ w ramach Cruces Mendel zauważył, że wzór dziedziczenia, który charakteryzował allele, które zdefiniowały cechę, nie wpłynęło na wzór dziedziczenia alleli, które zdefiniowały do innej cechy.
[TOC]
Teorie dziedziczenia postmendelskiego
Ponieważ Gregor Mendel zaproponował swoje słynne prawa ponad 100 lat nowych mechanizmów zostało wyjaśnione w celu wyjaśnienia dziedziczenia postaci, z których wiele można powiedzieć, że jest sprzeczne lub różni się znacznie od tych opisanych przez Mendla.
Przykładem zjawisk opisanych przez „współczesną” naukę, której postępy były możliwe tylko dzięki postępowi eksperymentalnych strategii i metod badawczych, są między innymi dziedzictwo związane z seksem, niepełną dominacją i kodominacją.
- Dziedzictwo związane z seksem
Dziedziczenie związane z seksem ma związek z genami występującymi tylko w chromosomach seksualnych. U ludzi seks jest określany przez chromosomy X i Y. Kobiety mają dwie kopie X (xx) i mężczyzn jednego z X i drugiego Y (XY).
Może ci służyć: połączone geny Dziedzictwo powiązane z matką przez (Źródło: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licencje/nabrzeże/4.0) Via Wikimedia Commons)
Dziedzictwo powiązane z matką przez (Źródło: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licencje/nabrzeże/4.0) Via Wikimedia Commons) Geny znalezione w każdym z tych dwóch chromosomów to geny, które przyczyniają się do tych cech związanych z płcią. Geny na chromosomie X są obecne u mężczyzn i kobiet, ale geny i są tylko u mężczyzn.
Dziedziczenie związane z seksem zostało zaobserwowane i po raz pierwszy opisane przez Williama Batesona, jednego z pierwszych naukowców w „odkrywaniu” pracy Mendla i wykorzystaniu ich do przeprowadzania nowych eksperymentów na podstawie ich wniosków.
Bateson dokonał eksperymentalnych krzyżowców kurczaków z upieczami o różnych kolorach, niektóre z ciemnymi i gładkimi piórami, a inne z gładkimi piórami i wzorem w paski.
Przez te krzyże zauważył, że u potomstwa powstałe samce zawsze mieli upierzenia ze wzorem w paski.
Dopiero w trzeciej generacji, gdy mężczyzna porysowany z ciemnymi i gładkimi piórami, uzyskano niektóre koguty z czarnym upierzeniem. Ta proporcja wynikająca z krzyżówek zakwestionowała proporcje zaproponowane przez Mendla.
Dziedzictwo powiązane z x
Po pracy Batesona Thomas Henry Morgan znalazł logiczne wyjaśnienie tak dziwnych wyników uzyskanych przez Batesona, przypisując różnicę zabarwienia kurczaków, do której wzór pasiasty był zakodowany przez dominujący allel, który był na chromosomie X.
U kurcząt, w przeciwieństwie do człowieka, płeć heterogametyczna (xy) odpowiada kobietom, tymczasem płeć homogamiczna odpowiada mężczyznom (xx).
Allel badany przez Batesona i Morgana został następnie wyrażony przez wszystkie te samce o ślinie w paski, ale gdy mężczyzna odziedziczył dwa chromosomy X z allelem odpowiadającym czarnym zabarwieniu, potem pojawili się mężczyźni ciemnego upierzenia i bez porysowanego wzoru.
- Niekompletna dominacja
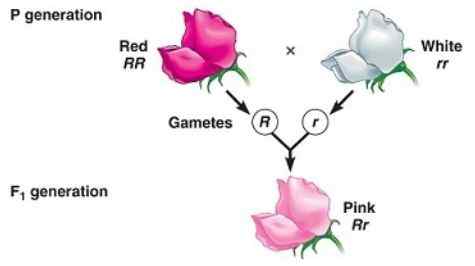 Pośredni fenotyp w pokoleniu F1 spowodowany niepełną dominacją
Pośredni fenotyp w pokoleniu F1 spowodowany niepełną dominacją Niekompletna dominacja ma miejsce, gdy fenotyp obserwowany u potomstwa, który wynika z przekroczenia dwóch homozygotycznych osobników (jeden dominujący i inny recesywny) jest fenotypem „pośrednim” między dominującym a recesywnym.
Ma to związek z „niezdolnością” „dominującego” allelu całkowicie maskowania ekspresji allelu recesywnego, dlatego wynikowy fenotyp odpowiada mieszance cech kodowanych przez oba allele, często opisywane jako „rozcieńczony fenotyp „Z dominującego.
Może ci służyć: monosomieNa przykład w nocy -latach (Mirabilis Jalapa) Typowy kolor płatków dzikich kwiatów jest czerwony. Kiedy roślina z czerwonymi płatkami przecina się z czystą rośliną z białymi płatkami, uzyskuje się, że wszystkie potomstwo to różowe płatki (różowe).
- Codominance
Kiedy dwa dominujące osoby homozygotyczne są skrzyżowane, aw potomstwie istnieje trzeci fenotyp, który odpowiada dwóch cech określonych przez dwa allele rodzicielskie, mówimy o Codominance.
W tym przypadku, zgodnie z przykładem podobnym do kwiatów poprzedniego przykładu, „trzeci fenotyp” nie odpowiadałby różowym kwiatom, ale roślinie z białymi kwiatami i czerwonymi kwiatami w tym samym łodydze, od dwóch alleli ” co -cake ”, a żaden nie maskuje drugiego.
Jednak wiele razy niekompletne kodominacja i dominacja są trudne do rozróżnienia fenotypowo, ponieważ oba wytwarzają własny fenotyp jednostki, który wydaje się wyrażać dwa różne allele.
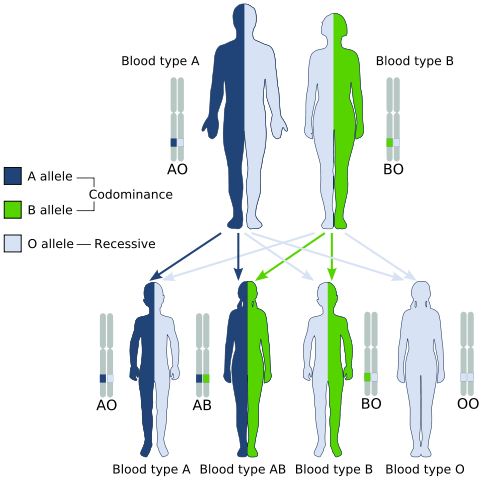 Codominance, przykład rodzaju dziedziczenia postmendelicznego (źródło: gyssinemRabettalk✉ Ten obraz z utworzeniem w 3C został utworzony z Inkscape. / Domena publiczna, przez wikimedia Commons)
Codominance, przykład rodzaju dziedziczenia postmendelicznego (źródło: gyssinemRabettalk✉ Ten obraz z utworzeniem w 3C został utworzony z Inkscape. / Domena publiczna, przez wikimedia Commons) Jednak genotypowo obecne osoby.
Klasycznym przykładem Codominance jest rodzaj grupy krwi u ludzi, ponieważ allele, które kodują antygeny A i B w populacjach ludzkich.
To znaczy, gdy allele A i B znajdują się razem w komórkach jednostki, ma typ AB krwi.
- Śmiercionośne allele
Są to recesywne allele dla jakiejś postaci, które, gdy są homozygotyczne, są śmiertelne dla jednostki. Osoby, które prezentują te allele, nie rodzą się, ponieważ unikają normalnego funkcjonowania maszyn molekularnych i unikają tłumaczenia genów.
- Interakcje genowe
Wiele miejsc w genomie indywidualnej oddziałują z innymi częściami tego samego genomu, to znaczy, że punktualna modyfikacja allelu wewnątrz genomu może bezpośrednio warunkować ekspresję innych alleli w komórkach jednostki.
Może ci służyć: wpływ inżynierii genetycznej na społeczny, ekonomiczny i środowiskowyTe interakcje są trudne do ustalenia i są podzielone na dwa różne typy: geny, które oddziałują na tej samej trasie genetycznej i te, które robią to na różnych trasach genetycznych.
Interakcje na różnych trasach genetycznych
Klasyczny i znany przykład genów, które oddziałują na różnych drogach genetycznych, ma związek z genami, które określają kolor skóry węży kukurydzianych (Panterophis guttatus), którego wspólnym kolorem jest powtarzalny wzór czarnej i pomarańczowej plamki.
 Wąż kukurydziany (źródło: factumquintus, przez Wikimedia Commons)
Wąż kukurydziany (źródło: factumquintus, przez Wikimedia Commons) Ten fenotyp jest wytwarzany przez dwa różne pigmenty, które są pod ścisłą kontrolą genetyczną. Gen określa pomarańczę i ma dwa allele: jeden lub+, który koduje pigment i drugi O-, który określa całkowity brak pigmentu pomarańczowego.
Kolejny inny gen określa obecność lub brak czarnego pigmentu, allele tego genu są znane jako B+, ten, który określa obecność czarnego pigmentu i B-, co określa brak pigmentu.
Oba geny nie są powiązane i określone niezależnie kolor węży, to znaczy wąż z genotypem lub+/b+ jest normalnym węża z fenotypem pomarańczowym i czarnym, ale każda zmienność genotypowa spowoduje inny fenotyp.
Interakcje na tej samej trasie genetycznej
Klasycznym przykładem tych genów jest opisany dla kwiatów „dzwonków” (z rodziny Campanulae), których „dziki” fenotyp odpowiada roślinom z niebieskimi kwiatami.
Niebieski kolor kwiatów dzwonowych jest określany nie przez gen, ale trasą metaboliczną, w której kilka enzymów, każde zakodowane przez inny gen, uczestniczy w syntezie pigmentu.
Zatem istnieją fenotypy charakteryzujące się wynikiem interakcji kilku genów, których produkty uczestniczą w tej samej trasie metabolicznej i gdzie mutacja lub zła funkcja któregokolwiek z nich, daje fenotyp, w tym przypadku białych kwiatów, deponowanych.
Bibliografia
- Ceccalli, zm. (2019). Historia dziedziczenia.
- Griffiths, s. 1., I Stotz, K. (2013). Genetyka i filozofia: wstęp. Cambridge University Press.
- Lewin, ur., Krebs, J. I., Goldstein, e. S., & Kilpatrick, s. T. (2014). Geny Lewina xi. Jones & Bartlett Publishers.
- McCleran, g. I., Vogler, g. P., & Plomin, r. (1996). Genetyka i medycyna behawioralna. Medycyna behawioralna, 22(3), 93-102.
- Richmond, m. L. (2008). Program badawczy Williama Batesona przed i postmendelijskim w „Dziedziczności i rozwoju.". Kulturalna historia dziedziczności IV: dziedziczność w stuleciu genu, 343, 213-242.
- « Zasady drażliwości komórkowej, składniki, odpowiedzi (przykłady)
- Wymarły i aktualny megafauna (z przykładami) »