

Fosfatydilenololoamina
- 3815
- 680
- Eliasz Dubiel
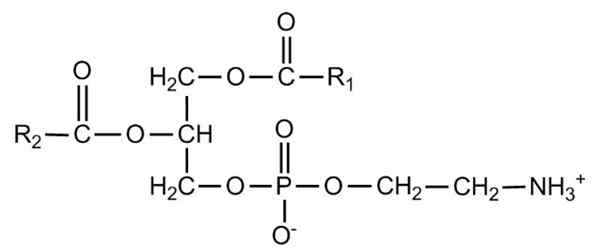
Półpośrednia formuła fosfatydyletanoloaminy. Źródło: Zirgouflex, Wikimedia Commons Co to jest fosfatydilenoloamina?
Fosfatydilenololoamina (PE) jest obfitym glyofosfolipidem w błonach komórkowych ludzkich tkanek. Pomimo jego obfitości zależy nie tylko od typu komórki, ale także od przedziału i czasu rozważanego cyklu życia komórek.
Błony biologiczne są barierami definiującymi organizmy komórkowe. Mają nie tylko funkcje ochrony i izolacji, ale są również kluczem do ustanowienia białek, które wymagają środowiska hydrofobowego dla optymalnego funkcjonowania.
Zarówno eukariot i prokaryot.
Glyceofosfolipidy to strukturalne cząsteczki amfipatyczne na szkielecie L-glicerolu, który jest estryfikowany w pozycjach SN-1 i SN-2 przez dwa kwasy tłuszczowe o długości i stopniu zmiennych nasycenia. W hydroksylu pozycji SN-3 jest esteryfikowana przez grupę fosforanową, która z kolei może łączyć różne rodzaje cząsteczek, które powodują różne rodzaje glicerofosfolipidów.
W świecie komórki istnieje wiele różnych gliofosfolipidów, jednak najliczniejsze są fosfatydylocholina (PC), fosfatydyletanoloamina (PE), fosfatydyloseryna (PS), fosfatydyloinitol (PI), fosfatyd (PA), fosfatydylooglysynool (pg) i karniolipina (PI), fosfatylinitolu (PI) (CL).
Struktura
Struktura fosfatydilenololaminy została odkryta przez Baer i współpracowników w 1952 roku. Jak określono eksperymentalnie dla wszystkich glicerofosfolipidów, fosfatydyletanoloamina powstaje przez cząsteczkę glicerolu sterylizowaną w pozycjach SN-1 i SN-2 z łańcuchami kwasów tłuszczowych między 16 a 20 atomami węgla.
Sterowane kwasy tłuszczowe w hydroksylu SN-1 są na ogół nasycone (bez podwójnych wiązań) o maksymalnej długości 18 atomów węgla, podczas gdy zjednoczone łańcuchy w pozycji SN-2 mają większą długość i z jednym lub więcej nienaturalnym (podwójne połączenia).
Może ci służyć: krążenie grzybów: składniki odżywcze, substancje, osmoregulacjaStopień nasycenia tych łańcuchów przyczynia się do elastyczności błony, co ma duży wpływ na wprowadzenie i porwanie białka w dwulicznicy.
Fosfatydyletanoloamina jest uważana za niehamellar glyceofosfolipid, ponieważ ma stożkowy kształt geometryczny. Ten kształt jest dany przez niewielki rozmiar grupy polarnej lub „głowy”, w odniesieniu do kształtu łańcuchów kwasów tłuszczowych, które zawierają hydrofobowe „kolejki”.
„Głowa” lub polarna grupa fosfatydyletanoloaminy jest znakiem ztolioniowym, to znaczy grupy, które mogą być pozytywne i negatywnie naładowane w pewnych warunkach pH.
Ta funkcja pozwala ustalić wiązania wodorowe z dużą ilością odpadów aminokwasowych, a ich rozkład obciążenia jest istotnym wyznacznikiem topologii domen wielu kompleksowych białek błonowych.
Biosynteza
W komórkach eukariotycznych synteza lipidów strukturalnych jest ograniczona geograficznie, jest głównym miejscem biosyntezy retikulum endoplazmatycznego (ER) i w mniejszym stopniu aparat Golgiego.
Istnieją cztery niezależne trasy biosyntetyczne do produkcji fosfatydyletanoloaminy: (1) trasa CDP-stanolaminy, znana również jako Kennedy Route. (2) Trasa PSD do dekarboksylacji fosfatydyloserynowej (PS). (3) Acylacja gładkich.
Trasa Kennedy
Biosynteza fosfatydyletanoloaminy tą drogą jest ograniczona do ER i wykazano, że w komórkach wątroby chomika jest to główna droga produkcji. Składa się z trzech kolejnych etapów enzymatycznych katalizowanych przez trzy różne enzymy.
Może ci służyć: flora i fauna Campeche: reprezentatywne gatunkiW pierwszym etapie występuje fosfoetanoloamina i ADP dzięki działaniu kinazy etanolaminy, która katalizuje fosforylację etanolaminy zależną od ATP.
W przeciwieństwie do roślin, ani ssaki, ani drożdże nie są w stanie wytwarzać tego substratu, więc należy je spożywać w diecie lub uzyskać z degradacji fosfatydyletalaminy lub sferycznych cząsteczek fosfatydilnych lub sphygozyny.
Fosfoetanoloamina jest wykorzystywana przez CTP: fosfoetanoloamina cytidiltransferaza (ET) w celu utworzenia CDP o wysokiej energii: etanoolamina i fosforan nieorganiczny.
1,2-dicyloglicerolu etanoolamina fosfotransferaza (ETP) wykorzystuje energię zawartą w łączu CDP-eneanolaminy z kowalencyjnie zjednoczającą etanolaminę z cząsteczką diacyloglicerolu.
Trasa PSD
Ta trasa działa zarówno w prokariotach, jak i drożdżach i ssakach. W bakteriach występuje w błonie plazmatycznej, ale w eukariotach ma miejsce w obszarze retikulum endoplazmatycznego, który ma ścisły związek z błoną mitochondrialną.
U ssaków trasa jest katalizowana przez pojedynczy enzym, dyskuarboksylaza fosfatydyloseryny (PSD1P), która jest osadzona w błonie mitochondrialnej, której gen jest kodowany przez jądro. Reakcja implikuje dekarboksylację PS na fosfatydyletanoloaminę.
Pozostałe dwie drogi (acylacja gładkiej PE i wymiana grupy polarnej zależna od wapnia) występują w retikulum endoplazmatycznym, ale nie przyczyniają się znacząco do całkowitej produkcji fosfatydilenoloaminy w komórkach eukariotycznych w komórkach eukariotycznych w eukariotycznych.
Funkcje
- Glyceofosfalipidy mają trzy główne funkcje w komórce, w tym funkcje strukturalne, energii i oznakowania komórek.
- Fosfatydilenoloamina jest związana z zakotwiczeniem, stabilizacją i składaniem wielu białek błonowych, a także niezbędnymi zmianami konformacyjnymi dla działania wielu enzymów.
- Istnieją eksperymentalne dowody, które proponują fosfatydyletanoloaminę jako kluczowe glyceofosfolipid w późnym etapie telofazy, podczas tworzenia pierścienia skurczowego i ustanowieniu framoplastu, która umożliwia podział błony dwóch komórek.
- Ma również ważną funkcję we wszystkich procesach fuzji i rozszczepienia (związek i separacja) błon zarówno retikulum endoplazmatycznego, jak i aparatu Golgiego. W I. coli Udowodniono, że fosfatydyletanoloamina jest niezbędna do prawidłowego składania i funkcji permase enzym laktozy, dlatego zasugerowano, że odgrywa rolę molekularnego „opiekuna”.
- Fosfatydilenoloamina jest głównym dawcą cząsteczki etanoloaminy niezbędnej do potranslacyjnej modyfikacji licznych białek, takich jak kotwice GPI.
- Ten glyceofosfolipid jest prekursorem licznych cząsteczek o aktywności enzymatycznej. Ponadto cząsteczki pochodzące z jego metabolizmu, a także diaclicerolu, kwasu fosfatydowego i niektórych kwasów tłuszczowych, mogą działać jako drugi posłańcy. Dodatkowo jest to ważny substrat do produkcji fosfatydylocholiny.
Bibliografia
- Gibellini, f., I Smith, T. K. Kennedy szlak Novo Synteza fosfatidyloetanoloaminy i fosfatydylocholiny. IUBMB Life.
- Harayama, t., & Riezman, H. Zrozumienie różnorodności składu lipidów błonowych. Nature przegląd biologii komórek molekularnych.