

Składnik łańcucha przenośnika elektronów, sekwencja, inhibitory
- 4457
- 1180
- Filip Augustyn
łańcuch przenośnika elektronów Składa się z zestawu cząsteczek białka i koenzymu w błonie. Jak sama nazwa wskazuje, odpowiada za transport elektronów z koenzymów NADH lub FADH2 do końcowego odbiornika, który wynosi O2 (tlen molekularny).
W tym procesie transportu energia uwalniana przez przenoszenie elektronów z koenzymów do tlenu molekularnego przez centra redoks zjednoczone z białkiem, jest związana z produkcją energii (ATP). Energia ta jest uzyskiwana dzięki gradientowi protonu generowanego w mitochondrialnej membranie wewnętrznej.
Ten system transportu składa się z różnych komponentów, które można znaleźć w co najmniej dwóch stanach utleniania. Każdy z nich jest skutecznie zmniejszony i reoksyd podczas ruchu elektronów z NADH lub FADH2 do O2.
Koenzymy NAD+ i FAD są zmniejszone w trasach utleniania kwasów tłuszczowych i cyklu kwasu cytrynowego w wyniku utleniania różnych substratów. Następnie te koenzymy są utleniane w elektronicznym łańcuchu transportu.
Tak że elektroniczny system transportu składa się z sekwencji reakcji redukcji tlenku, które są ze sobą połączone.
[TOC]
Składniki łańcucha
W zależności od rodzaju organizmu można zaobserwować od 3 do 6 składników stanowiących łańcuch przenośnika elektronów. Proces transportu elektronów i synteza ATP przez fosforylację oksydacyjną są procesami występującymi w błonie.
W przypadku komórek prokariotycznych (bakterie tlenowe) procesy te zdarzają się związane z błoną plazmatyczną. W komórkach eukariotów występuje w błonie mitochondrialnej, więc elementy transportu elektronów znajdują się w wewnętrznej części błony.
https: // giphy.COM/GIFS/HTKSBBADEQ9Z7CO5E
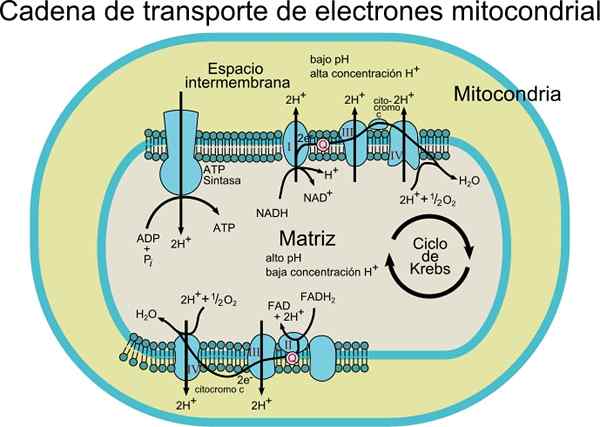
Elektrony są stopniowo przenoszone przez cztery kompleksy, które tworzą elektroniczny łańcuch transportu.
Każdy kompleks ma kilka składników białkowych związanych z grupami protetycznymi (składniki nieaminokwasowe białek koniugatowych) redoks, co pozwala na zwiększenie ich potencjałów redukcyjnych.
Ponadto ten system transportu składa się z różnych gatunków molekularnych, takich jak flawoproteina; koenzym, który zwany także ubiquinona (COQ lub UQ); różne cytochromy, takie jak cytochrom B, C, C1, A i A3; Białka z Fe-S i grupami białkowymi powiązanymi z Cu. Cząsteczki te są powiązane z błoną, z wyjątkiem cytochromu C.
Kompleks I
https: // giphy.com/gifs/rlf1rl9gdc6356sj6w
Kompleks I o nazwie Nadh Coenzyme quinona oksydortaduaza lub dehydrogenaza NADH, składa się z około 45 łańcuchów polipeptydowych i zawiera mononukleotydową cząsteczkę flawine (FMN) i od ośmiu do dziewięciu grup Fe-S Fe-S Fe-S Fe-S Fe-S Fe-S Fe-S (FMN). Jak sama nazwa wskazuje, ten kompleks przenosi parę elektronów z koenzymu NADH do COQ.
Funkcja kompleksu dehydrogenazy NADH zaczyna się od połączenia NADH do wspomnianego kompleksu po stronie wewnętrznej macierzy błony mitochondrialnej. Elektrony są następnie transportowane z NADH do FMN. Następnie elektrony przechodzą od zmniejszonej flawiny (FMNH2) do białek Fe-S.
FMNH2 działa jako rodzaj pomostu między białkami NADH i Fe-S, ponieważ ten ostatni może przenosić tylko pojedynczy elektron, podczas gdy koenzym NADH przenosi dwa, dzięki półcynon.
Może ci służyć: monocytopoiesis: etapy, cechy, regulacjaWreszcie elektrony są przenoszone z grup Fe-S do koenzymu Q, który jest mobilnym przenośnikiem elektronowym z izoprenedem ogonem, który sprawia, że jest hydrofobowy, umożliwiając przekroczenie środka membrany mitochondrialnej.
Kompleks II
![]()
Kompleks II, lepiej znany jako dehydrogenaza bursztynianu, jest integralnym białkiem mitochondrialnej błony wewnętrznej i jest enzymem, który interweniuje w cyklu kwasu cytrynowego.
Ten kompleks składa się z dwóch hydrofilowych podjednostek i dwóch hydrofobowych z grupami HEMO B, które zapewniają miejsce wiązania COQ, oprócz flawoproteiny i białka z Fe-S.
W cyklu kwasu cytrynowego (Krebs lub cykl kwasu trikarboksylowego), bursztynian przekształca się w fumaran przez dehydrogenazę bursztynianową, zmniejszając modę koenzymu do fadh2. Z tego ostatniego koenzymu elektrony są przenoszone do centrów Fe-S, które z kolei przenoszą je do COQ.
Podczas reakcji tego transferu elektronów standardowy potencjał redoks jest bardzo niski, co zapobiega uwalnianiu niezbędnej energii swobodnej do syntezy ATP.
Oznacza to, że kompleks II jest jedynym kompleksem w łańcuchu transportu elektronów, który nie jest w stanie zapewnić energii do syntezy ATP. Jednak ten kompleks jest kluczowy w tym procesie, ponieważ przenosi elektrony z FADH2 do reszty łańcucha.
Kompleks III
https: // giphy.com/gifs/cjg0raiuizjds2hkyu
Kompleks III, kompleks cytochromowy BC1 lub Reduktaza cytochromu C, przenosi elektrony z zmniejszonego koenzymu do cytochromu C. Ten transfer odbywa się przez pojedynczą drogę redoks, znaną jako cykl Q.
Ten kompleks składa się z białka z Fe-S i trzema różnymi cytochromami, w których atom żelaza zlokalizowany w grupie HEMO różni się cyklicznie między stanami zmniejszonymi (Fe2+) i utlenionym (Fe3+).
Cytochromy to hemoproteiny transportu elektronicznego, które mają aktywność redoks. Są obecne we wszystkich organizmach, z wyjątkiem niektórych obowiązkowych beztlewnych.
Białka te mają grupy hemo, które naprzemiennie między dwoma stanami utleniania (Fe2+ i Fe3+). Cytochrom C to mobilny transporter elektronów słabo związany z wewnętrzną błoną mitochondriów.
Cytochromy znalezione w tym kompleksie to cytochromy B, C i A, 3 są aktywnymi białkami redoks z grupami, mamy różne cechy, które naprzemiennie ich stany utleniania między Fe2+ i Fe3+.
Cytochrom C jest obwodowym białkiem błony, które działa jako „wahadło” elektronu z cytochromem C1 i z kompleksem IV.
IV kompleks
Cytochrom C i O2 są końcowymi receptorami elektronów pochodzących z utleniania materiału organicznego, więc kompleks oksydazy IV lub cytochromu C jest enzymem końcowym w procesie transportu elektronicznego. To akceptuje elektrony z cytochromu C i przenosi je do redukcji O2.
Złożona funkcja polega na katalizowaniu utleniania elektronu czterech kolejnych cząsteczek zmniejszonego cytochromu C, to znaczy jednocześnie zmniejsza cztery elektrony cząsteczki O2, ostatecznie wytwarzając dwie cząsteczki H2O.
Może ci służyć: tkanka powlekania nabłonkaSekwencja transportu elektronów
Elektrony są przenoszone z kompleksów I i II do kompleksu III dzięki koenzymie Q, a stamtąd przechodzą do kompleksu IV przez cytochrom C. Gdy elektrony przechodzą przez te cztery kompleksy, potencjał redukcji wzrasta, uwalniając energię, która jest następnie wykorzystywana do syntezy ATP.
W sumie przeniesienie kilku elektronów powoduje translokację 10 protonów przez membranę; cztery w kompleksach I i IV i dwa z kompleksu III.
Nadh descidrogenasa
Ten enzym katalizuje utlenianie koenzymu NADH przez koenzym, który. Elektrony przenoszą się z NADH do FMN, który jest przymocowany do hydrofilowego ogona kompleksu I. Grupowania Fe-S przenoszą się na elektrony. Te grupy Fe-S zmniejszają się do COQ, który jest osadzony w błonie, do ubikinolu (zmniejszony COQ).
Podczas przenoszenia elektronów do COQ cztery protony są z kolei przenoszone przez membranę wewnętrzną do przestrzeni międzyprzewodowej. Mechanizm, w którym translokowują się te protony, obejmuje białko znajdujące się w hydrofobowym ogonie kompleksu I.
Proces transferu elektronów w tym etapie uwalnia energię swobodną, w szczególności -16,6 kcal/mol.
Reduktaza COQ-cytokromo C i cykl Q
Koenzym, który jest utleniony przez cytochrom C, w reakcji katalizowanej przez ten koenzym. Utlenianie ubikinolu (zmniejszone COQ) występuje w pewnym miejscu kompleksu (qo lub miejsce utleniania) w błonie mitochondrialnej, przenosząc dwa elektron.
W cyklu Q utlenianie COQ wytwarza półchinon, w którym elektrony są przenoszone do grup HEMO B1 i BH. Podczas gdy występuje ten transfer elektronów, drugi COQ utlenia się w miejscu Qo, powtarzając cykl.
Cykl ten powoduje przeniesienie dwóch elektronów, a z kolei translokacja czterech protonów do przestrzeni międzybranowej, z uwalnianiem -10,64 kcal/mol energii swobodnej.
Citocroma C oksydaza
Ten enzym (kompleks IV) katalizuje utlenianie cytochromu C (zmniejszonego) przez O2, który jest akceptorem elektronów końcowych. Ten transfer wytwarza cząsteczkę H2O dla każdej pary elektronów przeniesionych oprócz translokacji protonów przez membranę.
Elektrony przenoszą się z jednego do jednego, z zmniejszonego cytochromu C do pary jonów CUA, a następnie przechodzą do hemo A i wreszcie docierają do centrum dwuklenowego kompleksu, które zawiera jony Cub i Hemo A3, gdzie przeniesienie czterech Elektrony występują w przypadku tlenu.
W kompleksie IV pierwiastki przenoszą elektrony jeden po drugim, tak że O2 jest stopniowo zmniejszone, tak że nie występuje uwalnianie niektórych toksycznych związków, takich jak nadtlenek, nadtlenek wodoru lub rodniki hydroksylowe.
Energia uwolniona na tym etapie odpowiada -32 kcal/mol. Gradient elektrochemiczny wygenerowany podczas procesu przenoszenia i zmiany energii (ΔE) spowodowane przez kilka elektronów podczas przechodzenia przez cztery kompleksy, na każdym etapie odpowiada swobodnej energii niezbędnej do produkcji cząsteczki ATP.
Może ci służyć: teoria składania membranyWielka dehydrogenaza
Jak wspomniano, ten kompleks ma jedyną, ale ważną funkcję wprowadzania elektronów FADH2 z cyklu kwasu cytrynowego do łańcucha transportu elektronów.
Ten enzym katalizuje utlenianie koenzymu FADH2 przez koenzym Q (utleniony). W cyklu kwasu cytrynowego, gdy bursztynian do fumaranu, dwa elektrony i dwa protony są przenoszone na modę. Następnie FADH2 przenosi te elektrony do COQ przez centra FE-S kompleksu.
Wreszcie, z COQ elektrony są przenoszone do kompleksu III, zgodnie z wyżej opisanymi krokami.
Kompleksy łańcuchowe są niezależne
Cztery kompleksy, które składają się na elektroniczny łańcuch transportu, są niezależne, to znaczy są niezależnie i działają w mitochkondrialnej membranie wewnętrznej, a ruch każdej z nich w błonie nie zależy lub jest powiązany z innymi kompleksami.
Kompleksy I i II poruszają się w membranie, przenosząc swoje elektrony do COQ, które rozprzestrzeniają się również w membranie i przenosi je do kompleksu III, skąd elektrony przechodzą do cytochromu C, który jest również mobilny w membranie i osadza elektrony w kompleksie IV złożonym.
Elektroniczne inhibitory łańcucha transportu
Na elektronicznym łańcuchu transportu niektóre specyficzne inhibitory działają w ich procesie. Rotenon jest powszechnie stosowanym środkiem owadobójczym, który jest stechiometrycznie połączony z kompleksem I, unikając zmniejszenia COQ.
Niektóre leki typu barbiturany, takie jak Piericidyna i Amytal, hamują kompleks I, zakłócając przenoszenie elektronów z grup Fe-S do COQ.
W kompleksie II niektóre związki, takie jak oiltrifluoroaceton i malonian, działają jako konkurencyjne inhibitory z bursztynian.
Niektóre antybiotyki, takie jak mixotiazol i stygmatellina, wiążą się z miejscem Q COQ, hamując przenoszenie elektronów z koenzymu Q do ośrodków Fe-S białek.
Cyjan, Azida (N3-), kwas siarkowy i tlenek węgla hamują kompleks IV. Związki te wiążą się z grupami HEMO, unikając przeniesienia elektronów do złożonego lub tlenu (O2) Binjądarowego (O2).
Gdy łańcuch przenośnika elektronów jest hamowany, wytwarzanie energii jest zatrzymywane z powodu fosforylacji oksydacyjnej, powodując poważne uszkodzenia, a nawet śmierć organizmu.
Bibliografia
- Alberts, ur., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, p. (2004). Niezbędna biologia komórki. Nowy Jork: Garland Science. 2. edycja.
- Cooper, g. M., Hausman, r. I. & Wright, n. (2010). Komórka. (PP. 397-402). Wyd. Marbán.
- Devlin, t. M. (1992). Podręcznik biochemii: z korelacjami klinicznymi. John Wiley & Sons, Inc.
- Garrett, r. H., I Grisham, C. M. (2008). Biochemia. Wyd. Thomson Brooks/Cole.
- Rawn, J. D. (1989). Biochemia (nie. 577.1 surowe). Wyd. Międzyamerykański-McGraw-Hill
- Voet, d., & Voet, J. G. (2006). Biochemia. Wyd. Pan -american Medical.
- « Charakterystyka anguili elektrycznej, siedlisko, reprodukcja
- Charakterystyka biozoos, morfologia, reprodukcja, odżywianie »