

Funkcje pompy wapniowej, typy, struktura i działanie
- 2739
- 184
- Estera Wojtkowiak
Pompa wapnia Jest to struktura natury białkowej, która jest odpowiedzialna za transport wapnia przez błony komórkowe. Ta struktura jest zależna od ATP i jest uważana za białko typu ATPASA, zwane także Ca2+-Atpasa.
CA2+-ATPASA znajduje się we wszystkich komórkach organizmów eukariotycznych i są niezbędne do homeostazy wapnia w komórce. To białko przeprowadza pierwotny aktywny transport, ponieważ ruch cząsteczek wapnia jest sprzeczny z jego gradientem stężenia.
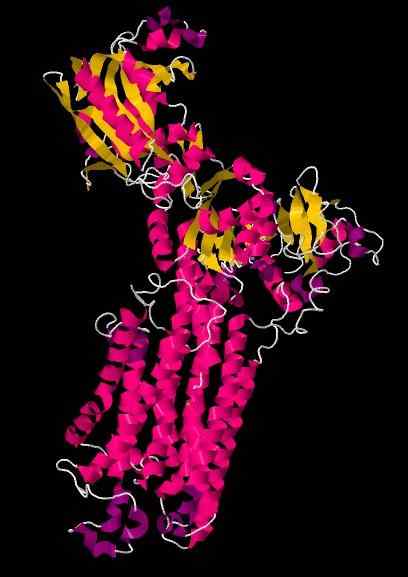
SERCA Struktura krystalograficzna.
Źródło: WCNSaffo [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] [TOC]
Funkcje pompy wapniowej
CA2+ Spełnia ważne artykuły w komórce, więc ich regulacja w nich ma fundamentalne znaczenie dla ich prawidłowego działania. Często działa jako drugi posłaniec.
W przestrzeniach pozakomórkowych stężenie CA2+ To jest około 10.000 razy wyższe niż w komórkach. Wzrost stężenia tego jonu w cytoplazmie komórkowej wywołuje różne odpowiedzi, takie jak skurcze mięśni, uwalnianie neuroprzekaźników i degradacja glikogenu.
Istnieje kilka sposobów przeniesienia tych jonów z komórek: transport pasywny (wyjście niespecyficzne), kanały jonowe (ruch na rzecz gradientu elektrochemicznego), antyportowy wtórny transport aktywny (NA/CA) i pierwotny transport aktywny z pompą ATP zależną.
W przeciwieństwie do innych mechanizmów przemieszczenia CA2+, Pompa działa w formie wektorowej. To znaczy, jon porusza się w jednym kierunku, aby działał tylko przez wydalanie.
Komórka jest wyjątkowo wrażliwa na zmiany stężenia CA2+. Przedstawiając tak wyraźną różnicę ze stężeniem zewnątrzkomórkowym, dlatego tak ważne jest skuteczne przywrócenie normalnych poziomów cytozolowych.
Może ci służyć: mesosomChłopaki
Opisano trzy rodzaje CA2+-Atasas w komórkach zwierzęcych, zgodnie z ich lokalizacjami w komórkach; Pompy znajdujące się w błonie plazmatycznej (PMCA), zlokalizowane w retikulum endoplazmatycznym i błonie jądrowej (SERCA), a także w błonie aparatu Golgiego (SPCA).
Bomby SPCA transportują również jony MN2+ które są kofaktorami różnych enzymów matrycy aparatu Golgiego.
Komórki drożdżowe, z innych organizmów eukariotingowych i komórek roślinnych mają inne rodzaje Ca2+-Bardzo szczególne atasas.
Struktura
Pompa PMCA
W błonie plazmatycznej znajdujemy aktywny transport antyporty2+ W komórkach spoczynkowych i aktywności. W większości komórek w spoczynku kierownik transportu wapnia na zewnątrz jest pompa PMCA.
Te białka składają się z około 1.200 aminokwasów i ma 10 segmentów transbłonowych. W cytosolu są 4 główne jednostki. Pierwsza jednostka zawiera grupę amino-końcową. Drugi ma podstawowe cechy, które pozwala fosfolipidom dołączyć do aktywatorów.
W trzeciej jednostce znajduje się kwas asparaginowy z funkcją katalityczną i „poniżej” tego pasma wiążącego fluoresceinę ISO, w domenie Unii ATP.
W czwartej jednostce znajduje się opanowanie Unii do Calmodulin, miejsc rozpoznawania niektórych kinaz (A i C) i pasmów wiążących CA2+ Alosteric.
Pompa SERCA
Pompy SERCA są w dużej ilości w retikulum sarkoplazmatycznym komórek mięśniowych, a ich aktywność jest związana z skurczem i relaksacją w cyklu ruchu mięśni. Jego funkcją jest transport CA2+ Od cytozolu komórki do matrycy retikulum.
Może ci służyć: podstawowa warstwy: Charakterystyka i funkcjeBiałka te składają się z pojedynczego łańcucha polipeptydowego z 10 domenami transmarketowymi. Jego struktura jest zasadniczo równa strukturze białek PMCA, ale różni się tym, że mają one tylko trzy jednostki w cytoplazmie, będąc miejscem aktywnym w trzeciej jednostce.
Działanie tego białka wymaga równowagi obciążeń podczas transportu jonów. Dwa Ca2+ (przez ATP zhydrolizowane) są przemieszczane z cytosolu do matrycy siatki, przeciwko bardzo wysokim gradientowi stężenia.
Ten transport występuje antyportycznie, ponieważ jednocześnie dwa h+ Są kierowane do cytosolu z matrycy.
Mechanizm operacyjny
Bomby SERCA
Mechanizm transportu jest podzielony na dwa stany E1 i E2. W E1 miejsca Unii, które stanowią wysokie powinowactwo do CA2+ Są skierowane do cytosolu. W E2 miejsca Unii są skierowane w kierunku światła retikulum, które stanowi niskie powinowactwo do CA2+. Dwa jony CA2+ Zjadaj się po przeniesieniu.
Podczas związku i przeniesienia CA2+, Występują zmiany konformacyjne, w tym otwarcie mdomeny białka, które jest w kierunku cytosolu. Jony są następnie łatwiej związane z dwoma witrynami Unii wspomnianej domeny.
Związek dwóch jonów CA2+ promuje szereg zmian strukturalnych w białku. Wśród nich obrót niektórych domen (domena a), która reorganizuje jednostki bombowe, umożliwiając otwarcie w kierunku matrycy retikulum w celu uwolnienia jonów, które są oddzielone dzięki zmniejszeniu powinowactwa w miejscach Unii.
Protony h+ i cząsteczki wody stabilizują miejsce Unii w CA2+, powodując ponowne obrócenie domeny do swojego pierwotnego stanu, zamykając dostęp do retikulum endoplazmatycznego.
Może ci służyć: cariocinesisPompy PMCA
Tego rodzaju pompy znajdują się we wszystkich komórkach eukariotycznych i są odpowiedzialne za wydalenie Ca2+ w kierunku przestrzeni pozakomórkowej, aby utrzymać jego stężenie w stabilnych komórkach.
W tym białku jon ca jest transportowany2+ Przez zhydrolizowany ATP. Transport jest regulowany przez poziomy białka kalmoduliny w cytoplazmie.
Poprzez zwiększenie stężenia CA2+ Wzrost poziomu cytozolowego, kalmoduliny, które wiążą się z jonami wapnia. Kompleks CA2+-Calmodulin, a następnie montaże miejsce bomb bomby PMCA. Istnieje zmiana konformacyjna w pompie, która umożliwia odsłonięcie otworu na przestrzeń pozakomórkową.
Jony wapnia są uwalniane, przywracając normalne poziomy wewnątrz komórki. W konsekwencji kompleks CA2+-Calmodulin to Desasambla, zwracając konformację pompy do pierwotnego stanu.
Bibliografia
- Brini, m., & Carafoli i. (2009). Pompy wapnia w zdrowiu i chorobie. Recenzje fizjologiczne, 89(4), 1341-1378.
- Carafoli, e., & Brini, m. (2000). Pompy wapnia: podstawy strukturalne i mechanizm transportu transbłonowego wapnia. Obecna opinia w biologii chemicznej, 4(2), 152-161.
- Devlin, t. M. (1992). Podręcznik biochemii: z korelacjami klinicznymi.
- Latorre, R. (Ed.). (1996). Biofizyka i fizjologia komórkowa. Uniwersytet Sevilla.
- Lodish, h., Darnell, J. I., Berk, a., Kaiser, c. DO., Krieger, m., Scott, m. P., & Matsudaira, p. (2008). Biologia komórek mięśniowych. Macmillan.
- Pocock, g., I Richards, C. D. (2005). Fizjologia ludzka: podstawa medycyny. Elsevier Hiszpania.
- Voet, d., & Voet, J. G. (2006). Biochemia. Wyd. Pan -american Medical.
- « Charakterystyka, taksonomia i klasyfikacja Eumetazoa
- Charakterystyka sosny kolumbijskiej, siedlisko, reprodukcja i zastosowania »