

Charakterystyka i funkcje tonoploro
- 1811
- 500
- Maksymilian Kępa
Toneplast Jest to termin stosowany w biologii do identyfikacji wewnętrznych błon wakuoli w komórkach roślinnych. Ton ma selektywną przepuszczalność i otacza wodę, jony i substancje rozpuszczone w próżniach.
Istnieją dokładne badania składu molekularnego tonu, ponieważ białka transportowe znajdujące się w tych błonach regulują wzrost roślin, stres na zasolenie i wysuszenie i podatność na patogeny.
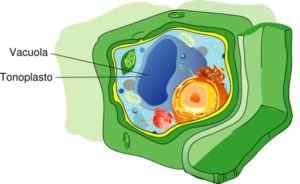 Ton komórki roślinnej (źródło: Mariana Ruiz [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons)
Ton komórki roślinnej (źródło: Mariana Ruiz [CC BY-SA 4.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons) Ogólnie rzecz biorąc, wakuola, którą komponuje ton, zawiera 57,2% całej objętości komórek w roślinach. Jednak odsetek ten może się różnić w zależności od sposobu życia, zwykle są rośliny kaktusa i pustynne, które mają wakuole o mniejszej lub większej wielkości.
U niektórych gatunków roślin wakuola ograniczona przez Toneplast może zajmować do 90% objętości wewnętrznej wszystkich komórek roślinnych.
Ponieważ bierze udział w stałym ruchu cząsteczek, jonów i enzymów między cytosolem a wnętrza wakuoli, ton jest bogaty w transport białek, kanałów i akwaporyn (pory lub kanały, w których przechodzi woda).
Wiele wewnętrznych pęcherzyków, takich jak fagosomy lub pęcherzyki transportowe.
Biotechnolodzy koncentrują swoje wysiłki na niezbędnych technikach włączenia, w roślinach o interesach komercyjnych, takich jak pszenica i ryż, ton z charakterystyką roślin odpornych na sól fizjologiczną.
[TOC]
Charakterystyka
Ton składa się głównie z białek i lipidów uporządkowanych w postaci biura lipidowej. Jednak w porównaniu z innymi błonami komórkowymi ma unikalne białka i lipidy w swoim składzie.
Membrana wakuolarna (ton) składa się z 18% neutralnych lipidów i steroli, 31% glikolipidów i 51% fosfolipidów. Zwykle kwasy tłuszczowe obecne w lipidach, które tworzą biway.
Może ci służyć: komórki diploidalneOgromna wakuola zdefiniowana przez Toneplast zaczyna się jako zestaw wielu małych wakuoli, które są syntetyzowane w retikulum endoplazmatycznym, wówczas białka z aparatu Golgiego są włączone.
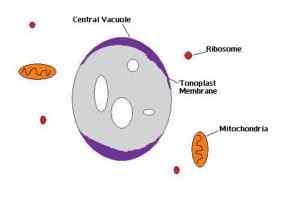 Schemat centralnej wakuoli komórki roślinnej (Źródło: Jestem autorem: Gevictor [CC BY-SA 3.0 (https: // creativeCommons.Org/licencje/by-sa/3.0)] przez Wikimedia Commons)
Schemat centralnej wakuoli komórki roślinnej (Źródło: Jestem autorem: Gevictor [CC BY-SA 3.0 (https: // creativeCommons.Org/licencje/by-sa/3.0)] przez Wikimedia Commons) Białka z aparatu Golgiego to kanały, enzymy, przenośniki i białka strukturalne oraz glikoproteiny kotwiczne, które będą pozycjonować ton ton.
Wszystkie małe wakuole łączą się i organizują powoli i stopniowo, tworząc ton, który powoduje dużą wakuola, głównie wypełnia wodę i jony. Proces ten występuje we wszystkich organizmach królestwa Plantae, Dlatego wszystkie komórki roślinne mają ton.
Ton, podobnie jak bituchondrialna biura lipidowa.
Funkcje
Główną funkcją tonu jest funkcjonowanie jako półprzepuszczalna bariera, wyznaczając przestrzeń rozumianą przez wakuola i oddzielając ją od reszty zawartości cytozolowej.
Ta „półprzestrzenność” jest wykorzystywana przez komórki roślinne pod kątem turgiczności, kontroli pH, wzrostu, wśród wielu innych funkcji.
Potencjał turgencji i wody
Najczęściej badaną funkcją roślin jest regulacja turgijności komórkowej. Stężenie jonów i wody, które znajdują się w wakuoli, uczestniczą, poprzez potencjał ciśnienia (ψp), w potencjalnym potencjale wodnym (ψ), tak aby cząsteczki wody dostały się lub opuszczają wewnątrz komórki.
Dzięki obecności tonu potencjał ciśnienia (ψp) wywierany jest protoplast (błona plazmatyczna) na ścianie komórkowej w komórkach. Siła ta nabiera dodatnie wartości, gdy wakuola wywiera nacisk na protoplast, a to z kolei na ścianę komórkową.
Kiedy woda opuszcza wakuola przez ton, a następnie opuszcza komórkę roślinną, wakuola zaczyna kurczyć się, a turgijność komórki jest tracona, osiągając wartości ciśnienia (ψp) bliskie zero, a nawet ujemne.
Może ci służyć: komórka Sertoli: Charakterystyka, histologia i funkcjeProces ten jest znany jako początkowa plazomoliza i to z kolei wytwarza więdnięcie, które obserwujemy w roślinach.
Gdy roślina jest połączona, jej potencjał osmotyczny (ψp) wzrasta, ponieważ gdy stężenie jonów potasowych (K+) wewnątrz komórki jest większe niż stężenie substancji rozpuszczonych na zewnątrz, woda porusza się do wewnątrz.
Te jony potasowe (K+) są głównie wewnątrz wakuoli i, dodane z jonami cytosolowymi, są odpowiedzialne za generowanie potencjału osmotycznego (ψp). Ton jest przepuszczalny dla tych jonów potasowych dzięki atpay, który ma w swojej strukturze.
Utrzymanie pH
Atasas w tonie.
ATP błony komórkowej korzeni są aktywowane przez obecność jonów potasowych (K+), wprowadzają one jony potasu (K+) i wydalenie protonów (H+). W przeciwieństwie do tego, atasy znajdujące się w tonie jest aktywowane w obecności chloru (Cl-) w cytosolu.
Te kontrolują stężenie jonów chloru (Cl-) i wewnętrznego wodoru (H+). Oba ATP działają w rodzaju „gry” w celu kontrolowania pH w cytosolu komórek roślin.
Gdy istnieje bardzo wysokie stężenie protonów (H+) w cytosolu, atass błony komórkowej wprowadza jony potasowe (K+); Podczas gdy atasa tonu jonów chloru jest do bani (Cl-) i wodoru (H+) cytosolu w wewnętrznej stronie wakuoli.
DOŁączenie jonów
Tonoplast ma kilka rodzajów pierwotnych pomp protonów. Ponadto ma kanały przenośne dla jonów wapnia (CA+), jonów wodoru (H+) i innych jonów, które są specyficzne dla każdego gatunku roślinnego.
Może ci służyć: egzocytoza: proces, typy, funkcje i przykładyAtasas pompowanie protonów (H+) w kierunku wnętrza wakuoli, dzięki czemu światło tego nabywa kwaśne pH, z wartościami między 2 a 5, a dodatnim obciążeniem częściowym. Pompy te hydrolizują ATP w cytosolu i, przez porów, wprowadzają protony (H+) w kierunku światła próżni.
Profosfaza to inny rodzaj „pomp” tonu, który również wprowadza protony (H+) do wakuoli, ale robią to poprzez hydrolizę pirofosforanu (PPI). Pompa ta jest wyłączna dla roślin i zależy od jonów MG ++ i K+.
W tonoplastu można znaleźć inne typy atasas, które pompują protony w kierunku cytosolu i wprowadzają jony wapnia (Ca ++) do wewnątrz próżni. Wapń (Ca ++) jest używany jako posłaniec w wnętrzu komórkowym, a światło wakuoli jest wykorzystywane jako depozyt tych jonów.
Być może najliczniejszymi białkami w tonie są kanały wapnia, które pozwalają one wyjście wapnia (CA+) wprowadzone przez membranowe atasas.
Obecnie zidentyfikowano również pierwotne bomby lub transportery typu ABC (z angielskiego DOTP-BInding CAsette) zdolne do wprowadzenia dużych jonów organicznych do wakuoli (na przykład glutationu).
Bibliografia
- Blumwald, e. (1987). Pęcherzyki TonePlast jako narzędzie w badaniu transportu jonów w próżni roślin. Physiologia Plantarum, 69 (4), 731-734.
- Dean, J. V., Mohammed, L. DO., & Fitzpatrick, T. (2005). Tworzenie, wakuolarna lokalizacja i toupoplast koniugatów glukozy kwasu salicylowego w hodowlach zawiesiny komórek tytoniowych. Roślina, 221 (2), 287-296.
- Gomez, L., & Chrispeels, m. J. (1993). Białka rozpuszczalne w tonie i wakuolarne są ukierunkowane przez różne mechanizmy. Komórka roślinna, 5 (9), 1113-1124.
- Jauh, g. I., Phillips, t. I., I Rogers, J. C. (1999). Tonoplast Wewnętrzne białko ISForms jako markery dla funkcji wakuolarnych. Komórka roślinna, 11 (10), 1867–1882.
- Liu, L. H., Ludewig, u., Gassert, ur., Frommer, w. B., I von Wirén, n. (2003). Transport mocznika przez regulowane azotem białka tonoplastowe w Arabidopsis. Fizjologia roślin, 133 (3), 1220-1228.
- Pesssarakli, m. (2014). Podręcznik fizjologii roślin i upraw. CRC Press.
- Taiz, L., Zeiger, e., Møller, i. M., & Murphy, a. (2015). Fizjologia i rozwój roślin