

Struktura receptorów muskarynowych, typy i ich funkcje, antagoniści
- 722
- 20
- Pani Gilbert Stolarczyk
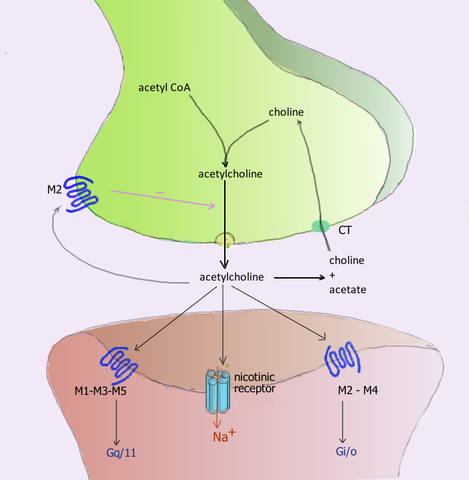
Receptory muskarynowe Są to cząsteczki, które pośredniczą w acetylocholinie (ACh) i znajdują się w postsynaptycznej błonie synapsów, w której uwalnia się wspomniany neuroprzekaźnik; Jego imię pochodzi od jego wrażliwości na alkaloid muscarynowy wyprodukowany przez grzyb Amanita Muscaria.
W ośrodkowym układzie nerwowym istnieje kilka zestawów neuronów, których aksony uwalniają acetylocholinę. Niektóre z nich kończą się w samym mózgu, podczas gdy większość tworzy szlaki motoryczne mięśni szkieletowych lub efektorowe ścieżki autonomicznego układu nerwowego dla gruczołów oraz serca i mięśni gładkich.
Neuroreceptor acetylocholiny podczas synapsy i ich odpowiednich receptorów w błonie postsynaptycznej (źródło: Użytkownik: Pancrat [GFDL 1.2 (http: // www.gnu antylopa.Org/licencje/stare licencje/FDL-1.2.html)] przez Wikimedia Commons) Acetylocholina uwalniana w związkach nerwowo -mięśniowych aktywnych receptorów cholinergicznych mięśni szkieletowych zwanych nikotynami, ze względu na jej wrażliwość na alkaloid nikotyny, które występują również w synapsach zwojów autonomicznego układu nerwowego (SNA).
Postoganglionowe neurony przywspółczulnego podziału tego układu wywierają ich funkcje poprzez uwalnianie acetylocholiny, które działają na muskarynowe receptory cholinergiczne znajdujące się w błonach komórków efektorowych, i indukując w nich modyfikacje elektryczne.
 Struktura chemiczna neuroprzekaźnika acetylocholiny (Źródło: Neurotokeker [domena publiczna] przez Wikimedia Commons) [TOC]
Struktura chemiczna neuroprzekaźnika acetylocholiny (Źródło: Neurotokeker [domena publiczna] przez Wikimedia Commons) [TOC]
Struktura
Receptory muskarynowe należą do rodziny receptorów metabotropowych, termin, z którymi receptory, które nie są odpowiednio kanałami jonowymi, są oznaczone, ale struktury białkowe, które po aktywowaniu wyzwalają wewnątrzkomórkowe procesy metaboliczne, które modyfikują aktywność prawdziwych kanałów.
Termin jest używany do odróżnienia ich od receptorów jonotropowych, które są prawdziwymi kanałami jonowymi, które otwierają się lub zbliżone przez bezpośrednie działanie neuroprzekaźnika, podobnie jak w przypadku receptorów nikotynowych wymienionych już z tabliczek nerwowo -mięśniowych mięśnia szkieletowego mięśnia szkieletowego.
W obrębie receptorów metabotropowych muskarynki są zawarte w grupie znanej jako Grozed G Aktywują fosfolipazę C (PLC).
Receptory muskarynowe są długimi kompleksowymi białkami błonowymi; Mają siedem segmentów transmarketowych złożonych z skłonności alfa, które sekwencyjnie przecinają dwuwarstwę błony lipidowej. Wewnątrz, na twarzy cytoplazmatycznej, są one związane z odpowiednim białkiem G, które transdukuje interakcję ligand-receiver.
Może ci służyć: Ewhriboblasts: Rozwój, funkcje i warstwyRodzaje receptorów muskarynowych i ich funkcje
Zidentyfikowano co najmniej 5 rodzajów receptorów muskarynowych, które są oznaczone za pomocą litery M, a następnie: M1, M2, M3, M4 i M5.
Receptory M1, M3 i M5 tworzą rodzinę M1 i charakteryzują się ich powiązaniem białka GQ lub G11, podczas gdy receptory M2 i M4 pochodzą z rodziny M2 i są związane z białkiem GI.
- Receptory M1
Występują głównie w ośrodkowym układzie nerwowym, w gruczołach zewnątrzopatrzenia i w zwojach autonomicznych Nodon. Są one sprzężone z białkiem GQ, które aktywuje enzym fosfolipazy C, który przekształca się w fosfatydylo inozytol (PIP2) w tryposforan inozytolu (IP3), który uwalnia wewnątrzkomórkowy, a dioplicerolu (DAG), który aktywuje białka C.
- Receptory M2
Znajdują się one zasadniczo w sercu, głównie w komórkach węzła chińskiego, na których działają one przez zmniejszenie częstotliwości rozładowania, jak opisano poniżej.
Automatyzm serca
Receptory M2 zostały dokładniej zbadane w węźle nodoaurykularnym (SA) serca, gdzie automatyzm, który okresowo wytwarza rytmiczne wzbudzenia odpowiedzialne za aktywność mechaniczną sercową, jest normalnie przejawowana.
Komórki węzła sinoaurykularnego, po każdym potencjale czynnościowym (PA) wyzwalającym skurcz sercowy (skurcz), są repolaryzowane i zwracane do poziomu około -70 mV. Ale napięcie nie pozostaje w tej wartości, ale cierpi postępująca depolaryzacja do poziomu progowego, który wywołuje nowy potencjał czynnościowy.
Ta postępująca depolaryzacja wynika z spontanicznych zmian prądów jonowych (i), które obejmują: K+ (IK1) redukcję wyjściową, pojawienie się prądu wejściowego Na+ (Fi), a następnie wejście Ca ++ (ICAT), aż osiągnie próg progowy a kolejny prąd Ca ++ (ICAL) odpowiedzialny za potencjał czynnościowy jest uruchamiany.
Jeśli wyjście K+ (IK1) jest bardzo niskie, a prądy wejściowe Na+ (if) i Ca ++ (ICAT) są wysokie, depolaryzacja występuje szybciej, potencjał czynności i skurcz występuje wcześniej, a seria częstotliwości jest wyższa. Przeciwne modyfikacje tych prądów niższa częstotliwość.
Może ci służyć: cariocinesisZmiany metabotropowe wywołane przez noradrenalinę (współczulną) i acetylocholinę (przywspółczulne) mogą zmienić te prądy. AMPC bezpośrednio aktywuje kanały IF, białka A (PKA) fosforyla i aktywuje kanały ICAT Ca ++, a grupa białek β GI aktywuje wyjście K+.
Działanie muskarynowe M2
Gdy acetylocholina uwalniana przez postanganglionowe zakończenia włókien błędnych serca (przywspółczulne) wiąże się z receptorami muscarynowymi M2 komórek węzła sinoaurykularnego, podjednostka αi białka GI zmienia jego GDP przez GDP i oddziela.
Podjednostka αi hamuje adenylcyklazę i zmniejsza wytwarzanie AMPC, co zmniejsza aktywność kanałów dla if i aktywność pKA. Ten ostatni fakt zmniejsza fosforylację i aktywność kanałów Ca ++ dla ICAT; Rezultatem jest zmniejszenie prądów depolaryzujących.
Grupa utworzona przez podjednostki βγ białka GI aktywuje prąd K+ Out (IKACH), który ma tendencję do przeciwdziałania wejściom Na+ i Ca ++ i obniża prędkość depolaryzacji.
Wynik stawu jest zmniejszenie nachylenia spontanicznej depolaryzacji i zmniejszenie częstości akcji serca.
- Receptory M3
 M3 Schemat receptora muskarynowego (źródło: Takuma-sa [CC0] przez Wikimedia Commons)
M3 Schemat receptora muskarynowego (źródło: Takuma-sa [CC0] przez Wikimedia Commons) Można je znaleźć w mięśniach gładkich (układ trawienny, pęcherz, naczynia krwionośne, oskrzela), w niektórych gruczołach zewnętrznych i w ośrodkowym układzie nerwowym.
Białko GQ są również sprzężone, a na poziomie płuc mogą powodować zwężenie oskrzeli, jednocześnie działając w naczyniowym śródbłonka.
- Receptory M4 i M5
Te receptory są mniej scharakteryzowane i badane niż poprzednie. Jego obecność w ośrodkowym układzie nerwowym i w niektórych tkankach obwodowych została zgłoszona, ale ich funkcje nie są jasno ustalone.
Może ci służyć: Tata Box: Charakterystyka i funkcjeAntagoniści
Uniwersalnym antagonistą tych receptorów jest atropina, alkaloid wyekstrahowany z rośliny Belladone Atropa, To wiąże się z nimi z wysokim powinowactwem, co stanowi kryterium odróżnienia od receptorów nikotynowych, które są niewrażliwe na tę cząsteczkę.
Istnieje wiele innych antagonistycznych substancji, które wiążą się z różnymi rodzajami receptorów muskarynowych o różnych powinowactwie. Połączenie różnych wartości powinowactwa dla niektórych z nich służyło dokładnie do włączenia tych receptorów do jednej z opisanych kategorii.
Częściowa lista innych antagonistów obejmowałaby: pirenzepinę, metotektraminę, 4-DAMP, hymbacinę, AF-DX 384, tripitaminę, darifenacynę, PD 102807, AQ RA 741, PFHHSID, MT3 i MT7; toksyny te ostatnie zawarte w truciznach odpowiednio zielonych i czarnych mabas.
Na przykład receptory M1 mają wysoką wrażliwość na pirenzepinę; M2 według tripitraminy, metctraminy i hybaciny; M3 przez 4-DAMP; M4 są bardzo powiązane z toksyną MT3, a także z hybaciną; M5 są bardzo podobne do M3, ale w odniesieniu do nich są mniej powiązane dla tutaj 741.
Bibliografia
- Ganong WF: Neurotransmitters i neuromodulatory, w: Przegląd fizjologii medycznej, 25. edycja. Nowy Jork, McGraw-Hill Education, 2016.
- González JC: Rola receptorów muskarynowych w modulacji transmisji gabaergicznej w hipokampie. Pamięć, aby zdecydować się na stopień doktora. Autonomiczny University of Madryt. 2013.
- Guyton AC, Hall JE: Rytmiczne wzbudzenie serca, w: Podręcznik fizjologii medycznej , 13. edycja; AC Guyton, Je Hall (red.). Philadelphia, Elsevier Inc., 2016.
- Piper HM: HerzeRregung, w: Physiologie des Menschen roztoczy patofizjologa, 31. edycja; RF Schmidt i in. (Eds). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, w: Fizjologia, 6. edycja; R Klinke i in. (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clafam DE, Schwartz JH: Modulacja transmisji synaptycznej: Second Messengers, In: Zasady nauk nerwowych, wydanie 5; E Kandel i in. (Eds). Nowy Jork, McGraw-Hill, 2013.
- « Historia biologii komórkowej, jakie badania, zastosowania i koncepcje
- Jak się rozmnażają rośliny? Rozmnażanie bezpłciowe i płciowe »