

Struktura włókien pośrednich, typy, funkcje
- 716
- 53
- Pani Waleria Marek
włókna pośrednie, Znany również w literaturze jako „IFS” (z angielskiego Pośrednie filmy), to rodzina nierozpuszczalnych cytozolowych białek włóknistych, które są obecne we wszystkich wielokomórkowych komórkach eukariotycznych.
Są częścią cytoszkieletu, który jest wewnątrzkomórkową siecią nitkowatnią, która jest głównie odpowiedzialna za wsparcie struktury komórkowej oraz różnych procesów metabolicznych i fizjologicznych, takich jak transport pęcherzyków, ruch i przemieszczenie komórek itp.
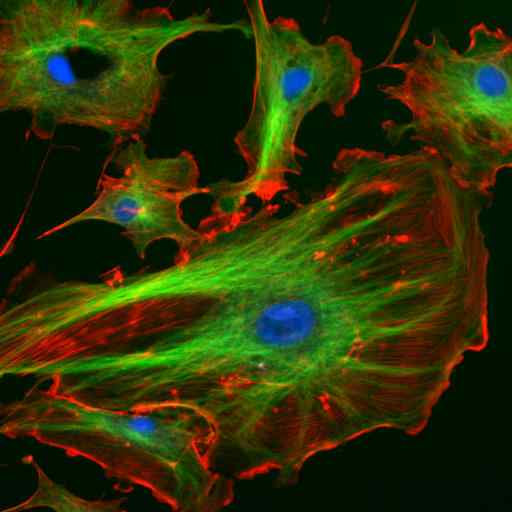
 Mikroskopia przez immunofluorescencję dwóch białek pośredniej włókien astrocytów (Vimentina i GFAP) (Źródło: Gerryshaw [CC BY-SA 3.0 (https: // creativeCommons.Org/licencje/by-sa/3.0)] przez Wikimedia Commons)
Mikroskopia przez immunofluorescencję dwóch białek pośredniej włókien astrocytów (Vimentina i GFAP) (Źródło: Gerryshaw [CC BY-SA 3.0 (https: // creativeCommons.Org/licencje/by-sa/3.0)] przez Wikimedia Commons) Wraz z mikrotubulami i mikrofilamentami, włókna pośrednie uczestniczą w przestrzennej organizacji organelli wewnątrzkomórkowych, w procesach endocytozy i egzocytozy, a także w procesach podziału komórek i komunikacji międzykomórkowej komunikacji.
Pierwszymi badanymi i opisanymi włóknami pośrednymi, były keratyny, jeden z pierwszych rodzajów białek, których struktura analizowano metodą dyfrakcji x -ray w latach 30. XX wieku.
Pojęcie włókien pośrednich zostało jednak wprowadzone w latach 80. XX wieku przez Lazarides, którzy opisali je jako „mechaniczne integratory przestrzeni komórkowej”, charakteryzujące się ich nierozwiązaniem i zdolnością do ponownego reensacji In vitro Po denaturacji.
Wielu autorów uważa się za „amortyzację” stresu dla komórek zwierzęcych, ponieważ są to bardziej elastyczne włókna niż mikrotubule i mikrofilamenty. Nie tylko występują w cytoszkielecie, ale są również częścią nukleoesqueleto.
W przeciwieństwie do innych włóknistych składników cytoszkieletu, włókna pośrednie nie uczestniczą bezpośrednio w procesach ruchliwości komórek, ale pracują w utrzymaniu strukturalnym i odporności mechanicznej komórek.
[TOC]
Struktura
Źródło: http: // rsb.Informacje.Nih.Gov/ij/Images/[domena publiczna] Włókna pośrednia mają przybliżoną średnicę 10 nm, strukturalną charakterystykę, którą zostały nazwane, ponieważ ich rozmiar jest między rozmiarami odpowiadającymi włókienom miozyny i aktyny, które wynoszą odpowiednio około 25 nm i 7 nm.
Różnią się one strukturalnie od pozostałych dwóch rodzajów włókien cytoszkieletowych, które są polimerami białkowymi kulisowymi, w których ich jednostki składowe są różnymi α-helikoidowymi białkami o dużej długości, które są grupowane od siebie, tworząc struktury w postaci ropy lub ropy.
Wszystkie białka, które składają się na włókna pośrednie, mają podobną organizację molekularną, która składa się z domeny α-heliksulidowej lub „linowej”, która ma różne segmenty „formatorów cewek” o tej samej wielkości.
Ta spiralna domena jest flankowana przez nieterminową „głowę” i niehypoidalny „ogon” na końcu C-końcowym i obie różnią się zarówno rozmiarem, jak i sekwencją aminokwasową.
W sekwencji tych dwóch skrajności znajdują się powody konsensusowe, które są wspólne dla 6 rodzajów znanych włókien pośrednich.
U kręgowców domena „liny” białka pośrednie.
Może ci służyć: gady: jakie są, cechy, klasyfikacja, reprodukcjaMontaż
Włókna pośrednie to struktury „samoorganiczne”, które nie mają aktywności enzymatycznej, która również odróżnia je od ich cytoszkieletowych odpowiedników (mikrotubule i mikrofilamenty).
Struktury te są początkowo montowane jako tetramery białek nitkowatych, które zawierają je pod wpływem tylko monowalentnych kationów.
Te tetramery mają o długości 62 nm, a ich monomery są ze sobą powiązane z boku, aby utworzyć „jednostki długości” (UFL, angielskiego Filmenty jednostkowe), Co jest znane jako faza 1 zespołu, która występuje bardzo szybko.
UFL są prekursorami długich włókien, a ponieważ dimery, które je zawierają, wiążą się w antyparalle i zatoczoną, jednostki te mają centralną domenę z dwiema domenami flankującymi, przez które występuje faza 2 wydłużenia, gdzie wzdłużne Unii Unii. innych UFL jest podawane.
Podczas tego, co nazywa się fazą 3 montażu, występuje zagęszczenie promieniowe średnicy włókien, co wytwarza dojrzałe włókna pośrednie o średnicy mniejszej lub mniejszej 10 nm.
Funkcje
Funkcje włókien pośrednich w znacznym stopniu zależą od rodzaju rozważanej komórki, a w przypadku zwierząt (w tym ludzi) ich ekspresja jest regulowana w sposób specyficzny dla tkanki, więc zależy to również od rodzaju tkanki niż w przypadku tkanki niż w przypadku tkanki niż w przypadku tkanki badanie.
Nabłonki, mięśnie, mezenchymalne i glejowe oraz neurony mają różne rodzaje włókien, specjalizowane według funkcji komórek, do której należą.
Wśród tych funkcji najważniejsze są strukturalne utrzymanie komórek i odporność na różne naprężenie mechaniczne, ponieważ struktury te mają pewną elastyczność, która pozwala im tłumić różne rodzaje sił narzuconych na komórki.
Rodzaje włókien pośrednich
Białka, które stanowią pośrednie włókna należą do dużej i heterogenicznej rodziny białek nitkowatych, które są chemicznie różne, ale rozróżniają się w sześciu klasach zgodnie z ich homologią sekwencji (I, II, III, IV, V i VI).
Chociaż nie jest to bardzo powszechne, różne typy komórek, w bardzo szczególnych warunkach (rozwój, transformacja komórek, wzrost itp.) Mogą współekspresować więcej niż jedną klasę białek tworzących włókno pośredniego
Klasa I i II włókna pośrednie: kwas i podstawowe keratyny
Keratyny reprezentują większość białek włókien pośrednich, a u ludzi reprezentują one ponad trzy czwarte włókien pośrednich.
Mają masę cząsteczkową, które różnią się od 40 do 70 kDa i różnią się od innych białek włókien pośrednich ze względu na ich wysoką zawartość glicyny i seryny.
Może ci służyć: jeże: cechy, siedlisko, reprodukcja, jedzenieSą znane jako kwas i podstawowe keratyny dla swoich punktów izoelektrycznych, które wynoszą między 4.9 i 5.4 dla kwaśnych keratyn i między 6.1 i 7.8 dla podstaw.
W tych dwóch klasach opisano około 30 białek i są obecne szczególnie w komórkach nabłonkowych, gdzie oba typy białka „współistnieją” i tworzą włókna złożone.
Wiele przypadków I Keratyny włókien pośrednich znajduje się w strukturach takich jak włosy, paznokcie, rogi, kolce i pazury, podczas gdy klasa II jest najliczniejsza w cytosolu.
Klasa III włókien pośrednich: białka typu demine/wimentyna
Falls jest kwaśnym białkiem o 53 kDa, które, w zależności od stopnia fosforylacji, ma różne warianty.
Niektórzy autorzy nazywali również włókna upadku jako „włókna pośrednie mięśni”, ponieważ ich obecność jest dość ograniczona, chociaż w małych ilościach wszystkie typy komórek mięśniowych.
W miofibrylach upadki występują w linii z, więc uważa się, że białko to przyczynia się do funkcji skurczowych włókien mięśniowych podczas pracy w połączeniu miofibrylii i błony plazmatycznej.
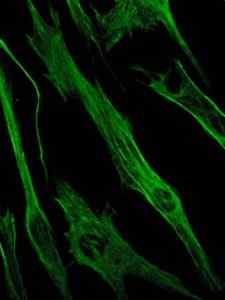 Zdjęcie barwienia białka wimentyny, białka pośrednie.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons)
Zdjęcie barwienia białka wimentyny, białka pośrednie.0 (https: // creativeCommons.Org/licencje/nabrzeże/4.0)] przez Wikimedia Commons) Z kolei wimentyna jest białkiem obecnym w komórkach mezenchymalnych. Pośrednie włókna utworzone przez to białko są elastyczne i ustalono, że są one odporne na wiele zmian konformacyjnych, które występują podczas cyklu komórkowego.
Występuje w fibroblastach, w komórkach mięśni gładkich, w białych krwinkach i innych komórkach układu krążenia zwierzęcego.
Klasa IV pośredniego włókien: białka neurorofilamentowe
Tego rodzaju pośrednie włókna, znane również jako „neurofilamenty”, obejmuje jeden z podstawowych elementów strukturalnych aksonów i dendrytów neuronalnych; Często są one związane z mikrotubulami, które również tworzą te struktury.
Neurorofilamenty zwierząt kręgowców zostały wyizolowane, określając, że jest to triplet białka 200, 150 i 68 kDa, który uczestniczy w montażu In vitro.
Różnią się od innych włókien pośrednich, w których mają ramiona poboczne, takie jak „dodatki”, które są rzutowane z ich peryferii i działają w interakcji między sąsiednimi włókienami a innymi strukturami.
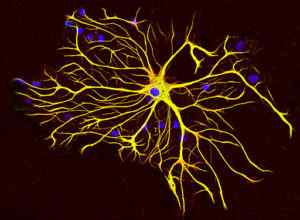
Komórki glejowe wytwarzają specjalny rodzaj włókien pośrednich znanych jako włókna pośrednie glejowe, które strukturalnie różnią się od neurofilamentów, ponieważ składają się z pojedynczego białka 51 kDa i które mają różne właściwości fizykochemiczne.
Klasa V włókna pośrednie: włókna blaszki jądrowej
Wszystkie arkusze, które są częścią nukleoesqueleto to w rzeczywistości pośrednie białka filamentu. Mają one od 60 do 75 kDa masę cząsteczkową i występują w jądrach wszystkich komórek eukariotycznych.
Może ci służyć: Trichuris TrichiuraSą one niezbędne dla wewnętrznej organizacji regionów nuklearnych i dla wielu funkcji tej niezbędnej organelli dla istnienia eukariotów.
Klasa VI włókien pośrednich: Nestines
Ten typ pośrednie włókna waży mniej więcej 200 kDa i znajduje się głównie w komórkach macierzystych ośrodkowego układu nerwowego. Są wyrażane podczas rozwoju neuronów.
Powiązane patologie
Istnieje wiele chorób u ludzi, które odnoszą się do włókien pośrednich.
Na przykład w niektórych rodzajach raka, takich jak złośliwe czerniaki lub rak piersi, na przykład koekspresja pośredniego włókien wimentyny i keratyny prowadzi do różnicowania lub interpretacji komórek nabłonkowych i mezenchymalnych.
Eksperymentalnie wykazano, że zjawisko to zwiększa aktywność migracyjną i inwazyjną komórek rakowych, co ma ważne implikacje w procesach przerzutowych charakterystycznych dla tego stanu.
Eriksson i współpracownicy (2009) przeglądają różne rodzaje chorób i ich związek ze specyficznymi mutacjami w genach zaangażowanych w tworzenie sześciu rodzajów włókien pośrednich.
Choroby związane z kodowaniem mutacji genów dla dwóch rodzajów keratyny są naskórka byka, naskórka, hiperkeratoza, dystrofia rogówki, rogowanie i wiele innych.
Włókna pośrednie typu III biorą udział w licznych kardiomiopatiach i różnych chorobach mięśni związanych głównie z dystrofią. Ponadto są również odpowiedzialne za dominujące zaćmę i niektóre rodzaje stwardnienia.
Liczne zespoły i zaburzenia neurologiczne są powiązane z włókienami typu IV, takimi jak Parkinson Parkinson. Podobnie wady genetyczne w włóknach typu V i VI są odpowiedzialne za rozwój różnych chorób autosomalnych i związane z funkcjonowaniem jądra komórkowego.
Przykładem są między innymi zespół Hutchinson-Gilford Progeria.
Bibliografia
- Andton, ur. H. (1981). Filamie pośrednie: rodzina homologicznych struktur. Journal of Muscle Research and Cell Motional, 2(2), 141-166.
- Eriksson, J. I., Pallari, h., Robert, zm., Eriksson, J. I., Dechat, t., Grin, ur.,… Goldman, r. D. (2009). Wprowadzenie Filamory pośrednie: Od odkrycia do choroby. Journal of Clinical Investigation, 119(7), 1763-1771.
- Fuchs, e., & Weber, k. (1994). Filamie pośrednie: struktura, dynamika, funkcja i choroba. Annu. Obrót silnika. Biochem., 63, 345-382.
- Hendrix, m. J. C., Seftor, e. DO., Chu i. W., Trevor, k. T., & Seftor, r. I. B. (1996). Rola wypełniaczy pośrednich w migracji, inwazji i przerzutach. Przegląd raka i przerzutów, piętnaście(4), 507-525.
- Herrmann, godz., & Aebi, u. (2004). Pracownice: struktura molekularna, mechanizm montażowy i integracja z funkcjonalnie wewnątrzkomórkowymi rusztowań wyraźne. Coroczny przegląd biochemii, 73(1), 749-789.
- Herrmann, godz., & Aebi, u. (2016). Filamy pośrednie: struktura i montaż. Perspektywy Cold Spring Harbor w biologii, 8, 1-22.
- McLean, ja., & Lane, b. (1995). Filamory pośrednie w Diew. Obecna opinia w biologii komórkowej, 7(1), 118-125.
- Steinert, s. 1., & Roop, D. (1988). Biologia molekularna i komórkowa filmów pośrednich. Coroczny przegląd biochemii, 57(1), 593-625.
- Steinert, s. 1., Jones, J., I Goldman, R. (1984). Pośrednie filmy. The Journal of Cell Biology, 99(1), 1-6.
- « Charakterystyka gasttrotryczna, siedlisko, reprodukcja, żywność
- Charakterystyka miękkiego meczetu, taksonomia, siedlisko, zastosowania »